Forget Typical Alzheimer's: AI Finds Four Types.
Quick Links
Is idiopathic Alzheimer’s more than one disease? And is there even such a thing as “typical” AD? Given the heterogeneity in clinical presentation and pathology, some scientists have suggested as much for years, but with the exception of certain rare, atypical forms, no consensus has emerged on subtypes of AD. That might be about to change. In the April 29 Nature Medicine, researchers led by Oskar Hansson at Lund University, Sweden, describe their analysis of the largest set of tau PET data to date. Their machine learning approach—an unbiased wrangling of thousands of scans from 1,667 people—groups AD into four distinct subtypes. Each has a different progression of neurofibrillary tangle pathology from region to region over time. Each also encompasses a distinct clinical profile and speed of worsening.
- Largest set of tau PET scans analyzed to date.
- Machine learning algorithm predicts four subtypes of AD.
- Diagnosis, prognosis, and monitoring of disease could change.
“Our data suggest the existence of multiple common AD subtypes, challenging the notion that there is such a pathological entity that can be described as ‘typical’ AD,” write the authors. Notably, even atypical forms of AD, such as posterior cortical atrophy and logopenic primary progressive aphasia, fit into this model as extreme forms of posterior and lateral temporal subtypes, respectively. In effect, the authors propose a single, unified model of AD progression.
Others found the work extremely exciting. “This is the kind of paper that can change a field,” Bernard Hanseeuw, Massachusetts General Hospital, told Alzforum. “We know that not all AD patients are equal and that they don’t have exactly the same set of symptoms or trajectories, but the power of their model is that they can explain both the symptoms and the progression of the disease.”
Ditto for Jonathan Schott, University College London. “It is a great paper with lots of important insights,” he wrote to Alzforum. “From a purely clinical standpoint, they question the existence of ‘typical' AD, reinforce the need for careful clinical phenotyping, and suggest that subtyping may in due course allow for more individualized approaches to patient care and prognostication,” he wrote (comment below).

Four Different Beasts. Based on tau PET scans, people with AD can be assigned to one of four AD trajectories. Each is different clinically. The older it starts, the milder the pathology. [Courtesy Vogel et al., 2021, Nature Medicine.]
First author Jacob Vogel, working in Alan Evans' lab at McGill University, Montreal, capitalized on a machine learning algorithm previously developed by co-author Alexandra Young at King’s College London. Young had used SuStaIn, short for Subtype and Stage Inference, to stitch together single MRI scans from almost 800 older volunteers. This identified three patterns of temporal and spatial atrophy in the brain, each with a different risk for developing AD (Oct 2018 news). In essence, SuStaIn extracted spatial patterns and longitudinal predictions from cross-sectional data.
“When I read Alexandra’s paper, I thought ‘Wow, if you have enough individuals to build a model, then you could just take one scan and find not only a person’s subtype, but how far along they are in the disease trajectory,’” Vogel said. But to do this for tau PET, there was a snag: Where to get the massive number of scans needed to create the model? “This is where Oskar’s diplomacy came in,” Vogel quipped.
Hansson collaborated with the leaders of five different cohorts who shared their PET scans. Data came from BioFinder-1 at Lund; the Yonsei University College of Medicine, Seoul, South Korea; Avid Radiopharmaceuticals, Philadelphia; the University of California, San Francisco; and from ADNI (Ossenkoppele et al., 2018; Cho et al., 2018; Pontecorvo et al., 2019; La Joie et al., 2020).
For the discovery sample, Vogel collated a total of 1,143 individual flortaucipir scans. Of these, 700 were negative for tangles and were designated subtype 0. From the remaining, positive scans, SuStaIn predicted that four subtypes best described the spatiotemporal patterns of neurofibrillary tangle progression (see image above). It also assigned each scan to one of 30 progressive stages along their respective subtype trajectories. Perhaps counterintuitively, severity negatively correlated with age, meaning that younger people tended to have more advanced pathology. This relationship has been seen before and hints that when disease strikes young, it tends to be more severe (Whitwell et al., 2019; Jack et al., 2020).
About 12 percent of the scans could not be categorized. These were of people who had very low tau burden and are likely cognitively normal. “Once people become cognitively impaired and deposit more tau, it is easier to subtype them,” said Vogel.
Of the remainder, about 33 percent of people were subtype 1. This might be closest to what researchers consider “typical” AD. S1 predicts a predominantly limbic pattern of tangle deposition with Braak-like spatial progression (see image below). Among the 18 percent of people categorized as S2, tangles predominantly emerge in the parietal lobe, with the medial temporal lobe being relatively spared. SuStaIn predicted 30 percent of individuals to be subtype 3. In them, tangles develop predominantly in the posterior occipital lobe and gradually expand toward the front of the brain. In 19 percent of the cohort who came out as subtype 4, tangles appear in the tempo-parietal cortices on the left side of the brain and gradually spread to parietal and frontal cortices.
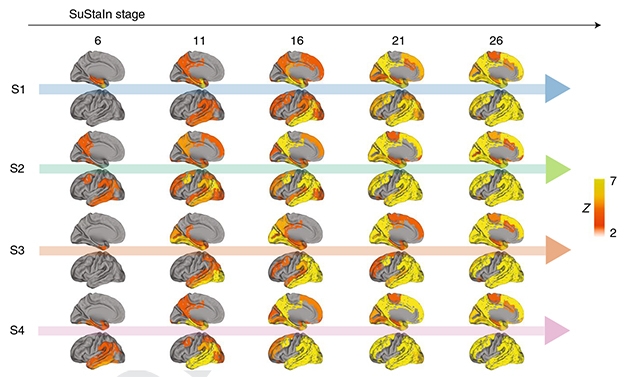
Subtypes and Stages. SuStaIn splits AD into four subtypes that progress over 30 stages. Each time point doublet shows the lateral and medial view of the left hemisphere. [Courtesy Vogel et al., 2021 Nature Medicine.]
Next, the authors ran SuStaIn on a replication sample comprising 469 people in BioFinder-2 who had been scanned with a different tau PET tracer, RO948. This time, they forced the machine learning algorithm to find four subtypes. Lo and behold, three of the four subtypes that it predicted highly resembled S1, S2, and S3. The spatiotemporal pattern of the fourth subtype looked like S4, but oddly, tangles were mostly in the right side of the brain, not the left. The authors believe the small sample size, not the tracer, drove this difference, and that the S4 subtype has a high propensity for either right or left dominance. This is a recognized feature of certain frontotemporal dementias, but not of Alzheimer’s.
How Do Subtypes Present in the Clinic?
Each subtype bar S3 had a symptom or demographic profile that distinguished it from the others. People with the S1 limbic subtype scored relatively better on global cognition, but worse on memory. They had less tangle burden than the other subtypes, and were more likely to carry an APOE4 allele.
The medial-temporal-sparing S2s, on the other hand, were less likely to carry APOE4. They were younger, had more tangles, and their executive function was weaker. The left-side-only S4s also tended to have more overall tau pathology, unsurprisingly mostly on the left sides of their brains. Though their memory was relatively preserved, they scored lower on overall cognition and language than the other groups. Language is typically processed in the left side of the brain. Women were less likely to be S4.

Slippery Subtype Slopes. Regardless of clinical diagnosis, the rate of decline on the MMSE was different for the four subtypes. People classified as S3 (orange) declined more slowly than those subtyped as S1 (blue), S2 (green) or S4 (purple). S4s worsened fastest. [Courtesy Vogel et al., 2021 Nature Medicine.]
A subset of 519 people from the initial five cohorts had follow-up scans. Among them, some subtype 0s had accumulated enough tangles between scans 1 and 2 to now be pinned as one of the four AD subtypes. Of the previously categorized S1-S4s, SuStaIn reclassified 84 percent the same way at their follow up, suggesting the staging was reproducible and stable. In support of this, those whose subtype did change had been categorized with low confidence on their first scan, underscoring the difficulty of assigning a subtype when a person's tau burden is still low.
As for clinical progression, MMSE scores for the S3s declined more slowly than for the other subtypes. This was true whether the person was cognitively normal at baseline or already had a diagnosis of mild cognitive impairment or AD. For S4s, it was the opposite. Their MMSE scores fell faster than any of the other subtypes.
SuStaIn stage also worsened faster in S4s, who declined about 1.73 stages per year. S1s and S3s declined at rates of 0.78 and 0.64 stages per year; for S2, it was a trend at 0.45 stages/year.
Could a clinician easily recognize any of these subtypes? “That would be tricky,” said Hansson. “S2 is the more cortical- and limbic-sparing, and people in this subtype tend to be younger, more dysexecutive, not as amnestic, and quite less likely to be ApoE4-positive. So if you see all that in the clinic you might think ‘S2,’ but for the others it would be difficult to predict.”
Why have these subtypes not popped out before? For one, the sheer number of scans this study analyzed is impressive, said Hanseeuw, and dwarfs the classic Braak pathological staging, which was based initially on 83 autopsy samples (Braak and Braak, 1991). For another, neuropathology is a patchy science. “Braak was limited in not being able to look at the whole brain, or use machine learning,” said Hansson. Typically, only one hemisphere of a brain donated for autopsy is used for pathology, which means one-sided effects are missed. Pathology is also constrained by spatial sampling and semi-quantitation. Even so, neuropathologists had identified hippocampal-sparing and limbic-predominant forms of AD, which correlated with different rates of atrophy on MRI (Murray et al., 2011; Whitwell et al., 2012). SuStaIn newly adds an unbiased look at whole-brain spatial and temporal variation.
What Does It All Mean?
Researchers see many opportunities for this proposed staging scheme to change the field. For his part, Hansson said, “Much more work is needed. First it’s important to find out how clinically relevant these subtypes are, for example to prognosis and progression rate. We need to study this in different cohorts.”
More broadly, future studies need to establish how these tangle-based subtypes relate to the many other aspects of Alzheimer's pathophysiology the field is exploring. For Eric Reiman, Banner Health, Phoenix, questions include, “for instance, what accounts for the different patterns of tangle deposition? To what extent does it impact a person’s phenotype, the relationship to CSF or plasma p-tau and to other biomarker measurements, their age at biomarker and clinical onset, their clinical course …” (see comment below).
What causes the different patterns is unclear. Could they be due to different conformations of toxic tau, i.e. seeds? Michel Goedert, Medical Research Council, Cambridge, U.K., suspects the seed is the same. Cryo-EM studies from his and other groups suggest that tau fibrils isolated from typical and atypical forms of AD are the same (Shi et al., 2021; July 2017 news; Falcon et al., 2018).
Or do the subtypes reflect how seeds propagate through the brain's connectome? On this idea, Hanseeuw thinks the subtypes may even depend on the way the brain has been wired from childhood. “It could be the same [tau] strain in each subtype, just on different tracks,” he said. Indeed, Vogel and colleagues noted that people with PCA and logopenic PPA are more likely to have particular learning disabilities in childhood, perhaps because of developmental differences (Feb 2008 news; Aug 2015 news).
Regarding future clinical trials, this study will have an impact, Hanseeuw believes. “It adds complexity. This is never easy to handle, but it is better to be aware of it, take it into account, and conduct better trials," he said. Others agreed. “For clinical trialists, the fact that different subtypes may have different clinical presentations, rates of progression, patterns of spread, and perhaps subtly different pathophysiology may lead to more bespoke clinical studies targeting not just different stages but different subgroups of AD,” Schott wrote. Reiman thinks this ability to recognize subtypes may affect the design and size of trials and boost the use of tau PET as an outcome measure.
Scientists might even reanalyze prior trials to learn how the four subtypes behaved. “If would be exciting, from a biological perspective, if a drug was more effective in one subtype,” said Vogel, “but there is a lot of work to be done. We hope others in the field will see if they can replicate our data, and use methods such as singe-cell sequencing to look for an explanation for the patterns.”—Tom Fagan
References
News Citations
- Across Time and Space: Machine Learning Reveals Paths to Dementia
- Tau Filaments from the Alzheimer’s Brain Revealed at Atomic Resolution
- Primary Progressive Aphasia—Learning Disabilities Point to Early Susceptibility
- Does Brain Development in Childhood Set the Stage for Dementia?
Paper Citations
- Ossenkoppele R, Rabinovici GD, Smith R, Cho H, Schöll M, Strandberg O, Palmqvist S, Mattsson N, Janelidze S, Santillo A, Ohlsson T, Jögi J, Tsai R, La Joie R, Kramer J, Boxer AL, Gorno-Tempini ML, Miller BL, Choi JY, Ryu YH, Lyoo CH, Hansson O. Discriminative Accuracy of [18F]flortaucipir Positron Emission Tomography for Alzheimer Disease vs Other Neurodegenerative Disorders. JAMA. 2018 Sep 18;320(11):1151-1162. PubMed.
- Cho H, Lee HS, Choi JY, Lee JH, Ryu YH, Lee MS, Lyoo CH. Predicted sequence of cortical tau and amyloid-β deposition in Alzheimer disease spectrum. Neurobiol Aging. 2018 Aug;68:76-84. Epub 2018 Apr 17 PubMed.
- Pontecorvo MJ, Devous MD, Kennedy I, Navitsky M, Lu M, Galante N, Salloway S, Doraiswamy PM, Southekal S, Arora AK, McGeehan A, Lim NC, Xiong H, Truocchio SP, Joshi AD, Shcherbinin S, Teske B, Fleisher AS, Mintun MA. A multicentre longitudinal study of flortaucipir (18F) in normal ageing, mild cognitive impairment and Alzheimer's disease dementia. Brain. 2019 Jun 1;142(6):1723-1735. PubMed.
- La Joie R, Visani AV, Baker SL, Brown JA, Bourakova V, Cha J, Chaudhary K, Edwards L, Iaccarino L, Janabi M, Lesman-Segev OH, Miller ZA, Perry DC, O'Neil JP, Pham J, Rojas JC, Rosen HJ, Seeley WW, Tsai RM, Miller BL, Jagust WJ, Rabinovici GD. Prospective longitudinal atrophy in Alzheimer's disease correlates with the intensity and topography of baseline tau-PET. Sci Transl Med. 2020 Jan 1;12(524) PubMed.
- Whitwell JL, Martin P, Graff-Radford J, Machulda MM, Senjem ML, Schwarz CG, Weigand SD, Spychalla AJ, Drubach DA, Jack CR Jr, Lowe VJ, Josephs KA. The role of age on tau PET uptake and gray matter atrophy in atypical Alzheimer's disease. Alzheimers Dement. 2019 May;15(5):675-685. Epub 2019 Mar 8 PubMed.
- Jack CR, Wiste HJ, Weigand SD, Therneau TM, Lowe VJ, Knopman DS, Botha H, Graff-Radford J, Jones DT, Ferman TJ, Boeve BF, Kantarci K, Vemuri P, Mielke MM, Whitwell J, Josephs K, Schwarz CG, Senjem ML, Gunter JL, Petersen RC. Predicting future rates of tau accumulation on PET. Brain. 2020 Oct 1;143(10):3136-3150. PubMed.
- Braak H, Braak E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991;82(4):239-59. PubMed.
- Murray ME, Graff-Radford NR, Ross OA, Petersen RC, Duara R, Dickson DW. Neuropathologically defined subtypes of Alzheimer's disease with distinct clinical characteristics: a retrospective study. Lancet Neurol. 2011 Sep;10(9):785-96. PubMed.
- Whitwell JL, Dickson DW, Murray ME, Weigand SD, Tosakulwong N, Senjem ML, Knopman DS, Boeve BF, Parisi JE, Petersen RC, Jack CR, Josephs KA. Neuroimaging correlates of pathologically defined subtypes of Alzheimer's disease: a case-control study. Lancet Neurol. 2012 Oct;11(10):868-77. PubMed.
- Shi Y, Murzin AG, Falcon B, Epstein A, Machin J, Tempest P, Newell KL, Vidal R, Garringer HJ, Sahara N, Higuchi M, Ghetti B, Jang MK, Scheres SH, Goedert M. Cryo-EM structures of tau filaments from Alzheimer's disease with PET ligand APN-1607. Acta Neuropathol. 2021 May;141(5):697-708. Epub 2021 Mar 16 PubMed. Correction.
- Falcon B, Zhang W, Murzin AG, Murshudov G, Garringer HJ, Vidal R, Crowther RA, Ghetti B, Scheres SH, Goedert M. Structures of filaments from Pick's disease reveal a novel tau protein fold. Nature. 2018 Sep;561(7721):137-140. Epub 2018 Aug 29 PubMed.
Further Reading
No Available Further Reading
Primary Papers
- Vogel JW, Young AL, Oxtoby NP, Smith R, Ossenkoppele R, Strandberg OT, La Joie R, Aksman LM, Grothe MJ, Iturria-Medina Y, Alzheimer’s Disease Neuroimaging Initiative, Pontecorvo MJ, Devous MD, Rabinovici GD, Alexander DC, Lyoo CH, Evans AC, Hansson O. Four distinct trajectories of tau deposition identified in Alzheimer's disease. Nat Med. 2021 May;27(5):871-881. Epub 2021 Apr 29 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University College London
To lump or to split? While Alzheimer’s disease is typically considered a late-onset, slowly progressive, amnestic syndrome, clinicians have long recognized substantial clinical heterogeneity. While in large part this may relate to the co-occurrence of other pathologies, including vascular disease, TDP43, and Lewy bodies (Boyle et al., 2018), even relatively “pure” AD may present with distinct phenotypes, including posterior cortical atrophy (the visual variant of AD), logopenic aphasia (early speech impairment), and rare cases of dysexecutive/behavioral AD and corticobasal syndrome AD (Graff-Radford et al., 2021). These variants, often overrepresented in young-onset cases, are associated with differential patterns of atrophy and tau deposition, and possibly also with different epidemiological, molecular, and genetic underpinnings (Graff-Radford et al., 2021).
Emerging evidence from pathological (Murray et al., 2011) and neuropsychology studies have shown, however, that while these clinical phenotypes may be extremes, even within so-called “typical AD” there is substantial heterogeneity. The advent of powerful computing techniques such as Subtype and Stage Inference (SuStaIn), has further advanced the ability to tease out subtypes in a data-driven manner. SuSTaIn was initially applied to MRI and neuropsychology data, parsing typical AD cases into three different subtypes (Young et al., 2018).
In this latest paper, Vogel and colleagues apply this powerful technique to a number of cohorts with tau PET data, providing evidence for the existence of four different subtypes—limbic, medial-temporal sparing, posterior, and lateral temporal variants—each with allied clinical and cognitive profiles, different longitudinal outcomes, and different predicted epicenters of tau deposition and spread (Vogel et al., Nature Medicine, in press). For those interested in atypical AD, the fact that the latter two subtypes appear to overlap with PCA and LPA, respectively, is of particular interest; but these intriguing findings have wide-ranging implications for the field of AD research more generally.
From a purely clinical standpoint they question the existence of “typical AD,” reinforce the need for careful clinical phenotyping, and suggest that subtyping may in due course allow for more individualized approaches to patient care and prognostication. From a basic science and clinical translational perspective, they demonstrate that simple one-size-fits-all staging models cannot capture the complexity of AD, suggesting instead a more complex scheme combining age, disease duration, and spatiotemporal tau distribution; defining and studying subtypes in this way may provide novel insights into pathological mechanisms including selective vulnerability and mechanisms of spread. For diagnostic frameworks, they reinforce the (known) limitations of categorizing individuals simply on the basis of presence/absence of AD pathologies—even before accounting for co-pathologies. And for clinical trialists the fact that different subtypes may have different clinical presentations, rates of progression, patterns of spread, and perhaps subtly differences in pathophysiology may lead to more bespoke clinical studies targeting not just different stages but different subgroups of AD.
References:
Boyle PA, Yu L, Wilson RS, Leurgans SE, Schneider JA, Bennett DA. Person-specific contribution of neuropathologies to cognitive loss in old age. Ann Neurol. 2018 Jan;83(1):74-83. Epub 2018 Jan 14 PubMed.
Graff-Radford J, Yong KX, Apostolova LG, Bouwman FH, Carrillo M, Dickerson BC, Rabinovici GD, Schott JM, Jones DT, Murray ME. New insights into atypical Alzheimer's disease in the era of biomarkers. Lancet Neurol. 2021 Mar;20(3):222-234. PubMed.
Murray ME, Graff-Radford NR, Ross OA, Petersen RC, Duara R, Dickson DW. Neuropathologically defined subtypes of Alzheimer's disease with distinct clinical characteristics: a retrospective study. Lancet Neurol. 2011 Sep;10(9):785-96. PubMed.
Young AL, Marinescu RV, Oxtoby NP, Bocchetta M, Yong K, Firth NC, Cash DM, Thomas DL, Dick KM, Cardoso J, van Swieten J, Borroni B, Galimberti D, Masellis M, Tartaglia MC, Rowe JB, Graff C, Tagliavini F, Frisoni GB, Laforce R Jr, Finger E, de Mendonça A, Sorbi S, Warren JD, Crutch S, Fox NC, Ourselin S, Schott JM, Rohrer JD, Alexander DC, Genetic FTD Initiative (GENFI), Alzheimer’s Disease Neuroimaging Initiative (ADNI). Uncovering the heterogeneity and temporal complexity of neurodegenerative diseases with Subtype and Stage Inference. Nat Commun. 2018 Oct 15;9(1):4273. PubMed.
Washington University
In this very nice work Vogel, Hansson, and colleagues found that in a large number of individuals with underlying AD pathology, four general patterns of tau pathology were observed based on tau-PET scans. These patterns were linked closely to the four types of clinical/anatomic patterns of cognitive decline seen in people who develop AD—1) amnestic/limbic; 2) non-amnestic cortical; 3) posterior cortical atrophy; and 4) logopenic aphasia.
Some previous pathological data, as well as more recently published clinical/tau PET data, have strongly suggested that these clinical patterns that are seen correlate well with the presence of tau pathology in specific areas of the brain that subserve the cognitive functions that decline. This large study strongly confirms and extends previous findings that these clinical patterns generally correspond well with tau accumulation in these different patterns, and it characterizes these patterns extremely well using advanced biostatistical techniques.
The findings have important implications for trying to better understand what leads to these different patterns mechanistically. In addition, in clinical trials, if they can be identified via both clinical and biomarker criteria with tau imaging, it will be important to determine if the different subtypes respond differently to different treatments.
Mayo Clinic
This is impressive work by Vogel and colleagues. It is very supportive of a line of investigation that we have been conducting that focuses on understanding individual variability in dementia using data-driven approaches coupled with detailed clinical phenotyping.
Using independent component analysis (ICA) of tau-PET scans, we found five patterns of tau deposition (Jones et al., 2017). Only one of these patterns was "typical"; the others were atypical and associated with younger age-of-onset, just as Vogel reports. One of the most striking patterns was nearly identical to a brain network associated with executive functioning (i.e., working memory) that we previously found in a data-driven analysis of functional MRI in 892 cognitively normal subjects in the Mayo Clinic Study of Aging (Jones et al., 2012).
After observing this network in functional MRI data, we began noticing this same pattern in FDG-PET scans of patients that we were seeing in our clinical practice. These individuals were younger than typical Alzheimer’s disease patients and had striking clinical features related to working memory dysfunction. They also did not have significant atrophy in the hippocampus. It became clear that these striking cases were a variation on a theme of predominate and early executive dysfunction as the defining clinical feature. We then defined the clinical construct of a progressive dysexecutive syndrome and collected our clinical experience with these cases in a recent manuscript (Townley et al., 2020). It should be emphasized that we focused only on individuals with biomarker evidence of AD, but other degenerative diseases can present with a progressive dysexecutive syndrome (Jones, 2020).
These individuals with positive AD biomarkers that met our criteria for a progressive dysexecutive syndrome had the exact same characteristics as the "S2 variant" described by Vogel and colleagues. They had less hippocampal atrophy, younger age-of-onset, were less likely to be ApoE4-positive, and multimodal imaging showed greater involvement of parieto-frontal brain regions on MRI, FDG-PET, and tau-PET. We also had detailed postmortem autopsy studies that showed that these cases had pathologically defined hippocampal sparing AD (Murray et al., 2011). We have also observed these relationships using hierarchical clustering of tau-PET and clinical-characteristic-based subcategorization (Lowe et al., 2018).
We also recently conducted a data-driven analysis of 492 patients on the AD spectrum, and have found that the imaging heterogeneity can be explained by a low-dimensional manifold we have termed the Global Functional State Space (Jones et al., 2020). The dysexecutive AD patients, or the "S2 variant," showed dysfunction in an extreme portion of the GFSS associated with executive functioning, further validating the clinical construct of dysexecutive AD.
We also found that all degenerative dementia syndromes exist along a continuum, specifically indexed by the GFSS, and distinct phenotypes occupy the extremes of this landscape.
References:
Jones D, Lowe V, Graff-Radford J, Botha H, Wiepert D, Murphy MC, Murray M, Senjem M, Gunter J, Wiste H, Boeve B, Knopman D, Petersen R, Jack C. Patterns of neurodegeneration in dementia reflect a global functional state space. medRxiv November 12, 2020. medRxiv.
Jones DT. Multiple aetiologies of the progressive dysexecutive syndrome and the importance of biomarkers. Brain Commun. 2020;2(2):fcaa127. Epub 2020 Nov 1 PubMed.
Jones DT, Graff-Radford J, Lowe VJ, Wiste HJ, Gunter JL, Senjem ML, Botha H, Kantarci K, Boeve BF, Knopman DS, Petersen RC, Jack CR Jr. Tau, amyloid, and cascading network failure across the Alzheimer's disease spectrum. Cortex. 2017 Dec;97:143-159. Epub 2017 Oct 3 PubMed.
Jones DT, Vemuri P, Murphy MC, Gunter JL, Senjem ML, Machulda MM, Przybelski SA, Gregg BE, Kantarci K, Knopman DS, Boeve BF, Petersen RC, Jack CR. Non-stationarity in the "resting brain's" modular architecture. PLoS One. 2012;7(6):e39731. PubMed.
Lowe VJ, Wiste HJ, Senjem ML, Weigand SD, Therneau TM, Boeve BF, Josephs KA, Fang P, Pandey MK, Murray ME, Kantarci K, Jones DT, Vemuri P, Graff-Radford J, Schwarz CG, Machulda MM, Mielke MM, Roberts RO, Knopman DS, Petersen RC, Jack CR Jr. Widespread brain tau and its association with ageing, Braak stage and Alzheimer's dementia. Brain. 2018 Jan 1;141(1):271-287. PubMed.
Murray ME, Graff-Radford NR, Ross OA, Petersen RC, Duara R, Dickson DW. Neuropathologically defined subtypes of Alzheimer's disease with distinct clinical characteristics: a retrospective study. Lancet Neurol. 2011 Sep;10(9):785-96. PubMed.
Townley RA, Graff-Radford J, Mantyh WG, Botha H, Polsinelli AJ, Przybelski SA, Machulda MM, Makhlouf AT, Senjem ML, Murray ME, Reichard RR, Savica R, Boeve BF, Drubach DA, Josephs KA, Knopman DS, Lowe VJ, Jack CR Jr, Petersen RC, Jones DT. Progressive dysexecutive syndrome due to Alzheimer's disease: a description of 55 cases and comparison to other phenotypes. Brain Commun. 2020;2(1):fcaa068. Epub 2020 May 27 PubMed.
Arizona Alzheimer's Consortium
This study capitalized on cross-sectional tau PET and related data from an extremely large number of research participants in preclinical, MCI, and dementia stages. It used a machine learning algorithm to characterize four spatial patterns of tau-tangle burden and explore the extent to which these patterns are related to the pattern and severity of cognitive characteristics, the presence or absence of the APOE4 allele, and demographic characteristics including sex. The authors suggest implications for clinical course and subsequent tau-tangle deposition. The study supports the suggestion from smaller tau PET studies that the pattern of tau-tangle deposition is more heterogeneous and individually variable than suggested by neuropsychological studies and Braak stage.
The study raises a number of intriguing questions that could be put to the test or confirmed in future studies. For instance, what accounts for the different patterns of tau-tangle deposition? To what extent does it impact a person’s phenotype? What is its relationship to CSF or plasma p-tau or other biomarker measurements, to age at biomarker and clinical onset or clinical course? How does it fit with pathophysiological progression, or differential response to disease-modifying treatments now in development? To what extent are these patterns related to APOE variants, other genetic and non-genetic risk and protective factors? What implications do the findings have for the size and design of clinical trials, participant enrichment or stratification, the analysis of tau PET images, and their role as a trial endpoint?
I congratulate the authors for their innovative and thoughtful efforts and this important contribution to the field, and I look forward to seeing ways in which the field can capitalize on the findings to further advance the fight against AD.
Northwestern University
The fact that Alzheimer’s disease (AD) is heterogeneous and that the Braak staging of neurofibrillary tauopathy can be violated has been known for nearly 30 years through work on posterior cortical atrophy and primary progressive aphasia (PPA) (Mesulam, 1982; Mesulam and Weintraub, 1992; Gefen et al., 2012; Rogalski et al., 2016; Hof et al., 1993; Hof et al., 1997; Jagust et al., 1990).
In approximately 40 percent of cases the progressive and relatively isolated language impairment of PPA is associated with AD neuropathology. When compared to the typical amnestic form of AD (DAT-AD), the form of AD that causes PPA (PPA-AD) is characterized by asymmetric cortical atrophy of the language-dominant (usually left) hemisphere, leftward predominance of neurofibrillary tangles (NFT), no association with ApoE4 as a risk factor, lesser frequency of limbic TDP-43, and closer linkage to familial dyslexia (Mesulam et al., 2021). In some cases, the language cortices have more NFT than limbic areas but in others it is the ratio of cortical-to-limbic NFT that is increased even when the medial temporal NFT density is at Braak stages 5-6. Only few PPA-AD cases are “hippocampal sparing” (Murray et al., 2011) postmortem, but most do have leftward asymmetry of NFT density (Gefen et al., 2012; Mesulam et al., 2021). What is remarkable is the preservation of memory in PPA-AD even when the postmortem shows typical NFT densities at Braak stages 5-6 (Mesulam et al., 2021).
The jaw-dropping 2,324 tau PET scans analyzed by Vogel et al. and the sophisticated SuStaIn analyses provide high-tech confirmation of heterogeneity in AD. The beauty of this approach is its potential ability to map tauopathy and therefore NFT distribution early in the course of the disease. However, while this obviates the problems of retrospective clinicopathologic inferences based on postmortem analyses, imaging modalities lack the precision of quantitative microscopy. It is also important to keep in mind that NFTs have complicated lifecycles. They emerge as intracellular paired helical filaments (PHFs), destroy the neurons within which they form, and remain as extracellular tombstones of lost neurons. Does the tau PET ligand penetrate the cell and identify nascent NFT at the PHF stage, or does it bind only the tombstones? A better understanding of the cellular correlates of PET ligands will enhance the value of information obtained through these spectacular modalities of molecular imaging.
References:
Mesulam MM. Slowly progressive aphasia without generalized dementia. Ann Neurol. 1982 Jun;11(6):592-8. PubMed.
Mesulam MM, Weintraub S, F. Boller et al., Editors. Primary progressive aphasia: Sharpening the focus on a clinical syndrome, in Heterogeneity of Alzheimer's Disease. Springer-Verlag: Berlin. 1992
Gefen T, Gasho K, Rademaker A, Lalehzari M, Weintraub S, Rogalski E, Wieneke C, Bigio E, Geula C, Mesulam MM. Clinically concordant variations of Alzheimer pathology in aphasic versus amnestic dementia. Brain. 2012 May;135(Pt 5):1554-65. PubMed.
Rogalski E, Sridhar J, Rader B, Martersteck A, Chen K, Cobia D, Thompson CK, Weintraub S, Bigio EH, Mesulam MM. Aphasic variant of Alzheimer disease: Clinical, anatomic, and genetic features. Neurology. 2016 Sep 27;87(13):1337-43. Epub 2016 Aug 26 PubMed.
Hof PR, Archin N, Osmand AP, Dougherty JH, Wells C, Bouras C, Morrison JH. Posterior cortical atrophy in Alzheimer's disease: analysis of a new case and re-evaluation of a historical report. Acta Neuropathol. 1993;86(3):215-23. PubMed.
Hof PR, Vogt BA, Bouras C, Morrison JH. Atypical form of Alzheimer's disease with prominent posterior cortical atrophy: a review of lesion distribution and circuit disconnection in cortical visual pathways. Vision Res. 1997 Dec;37(24):3609-25. PubMed.
Jagust WJ, Davies P, Tiller-Borcich JK, Reed BR. Focal Alzheimer's disease. Neurology. 1990 Jan;40(1):14-9. PubMed.
Mesulam M-, Coventry C, Bigio EH, Geula C, Thompson C, Bonakdarpour B, Gefen T, Rogalski EJ, Weintraub S. Nosology of Primary Progressive Aphasia and the Neuropathology of Language. Adv Exp Med Biol. 2021;1281:33-49. PubMed.
Murray ME, Graff-Radford NR, Ross OA, Petersen RC, Duara R, Dickson DW. Neuropathologically defined subtypes of Alzheimer's disease with distinct clinical characteristics: a retrospective study. Lancet Neurol. 2011 Sep;10(9):785-96. PubMed.
Mesulam MM, Coventry C, Kuang A, Bigio EH, Mao Q, Flanagan ME, Gefen T, Sridhar J, Geula C, Zhang H, Weintraub S, Rogalski EJ. Memory Resilience in Alzheimer Disease With Primary Progressive Aphasia. Neurology. 2021 Feb 9;96(6):e916-e925. Epub 2021 Jan 13 PubMed.
Institute of Clinical Neurobiology
Vogel et al. found four patterns of tau pathology based on tau PET scan in a large number of people. This pattern is similar to the one we proposed recently, suggesting four major subtypes of AD:
(1) Typical AD: frequency about 55 percent, balanced tau NFT counts in hippocampus and association cortex; multidomain amnestic syndrome; low final MMSE scores; high Aβ load in occipital cortex, less in middle temporal and parietal cortex; high burden of WMHs; frequent CAA; frequent APOE4 genotype, and increased Lewy and TDP-43 co-pathologies.
(2) Hippocampal-sparing AD: relative frequency 17 percent; tau NFT counts preliminary in neocortex; higher rate of cortical atrophy; young onset/death; male sex; nonamnestic syndrome; faster disease progression; most aggressive form; high cognitive reserve; APOE4 neg., low burden of WMHs; marked Aβ PET binding in frontal & parietal cortex; less frequent Lewy & TDP-43 co-pathologies.
(3) Limbic-predominant AD: relative frequency 21 percent; tau NFT counts predominantly in hippocampus; more severe atrophy in the medial temporal lobe; older age at onset/death; female sex; amnestic syndrome; slow disease progression; greater Aβ PET binding in frontal and parietal cortex; higher burden if WMHs; overloaded cognitive reserve; frequent APOE4 and MAPT H1H1 genotypes; hippocampal sclerosis; frequent TDP-43 co-pathology; less CAA, but more hypertensive angiopathy.
(4) Minimal atrophy AD: relative frequency 17 percent; no or minimal gray matter atrophy; intermediate age at onset; less intensive cortical tau pathology, but Aβ present; reduced metabolism in parietal cortex; low educational level/cognitive reserve; slow rate of progression; CAA may be present, but increased small vessel disease.
It is suggested that type 4 progresses to type 2 and further to type 1, as can type 3 progress to type 1, but type 2 can directly progress to type 1.
These four types appear not to be identical but in many respects similar to those described by Vogel et al. In addition, we should emphasize that only about 40-42 percent of all brains with AD pathology are "pure" AD types, while all the others are complicated by other co-pathologies.
References:
Jellinger KA. Pathobiological Subtypes of Alzheimer Disease. Dement Geriatr Cogn Disord. 2020;49(4):321-333. Epub 2021 Jan 11 PubMed.
Jellinger KA. Neuropathological assessment of the Alzheimer spectrum. J Neural Transm (Vienna). 2020 Sep;127(9):1229-1256. Epub 2020 Aug 1 PubMed.
Karolinska Institutet
Karolinska Institutet
We would like to congratulate Vogel and colleagues for their interesting article, which confirms, in a large tau PET dataset, the existence of four AD subtypes that are well known from neuropathological and MRI studies, as well as from clinical praxis.
In our systematic review from 2020, we foresaw that subtyping on tau PET would deliver important data to understand in-life trajectories of AD subtypes discovered at autopsy (Ferreira et al., 2020). During the last three years, several studies have discussed the topic, including one from our own lab, and now the study from Vogel and colleagues (Whitwell et al., 2018; Charil et al., 2019; Jeon et al., 2019; Ossenkopelle et al., 2020; Mohanty et al., 2020). A common thread in all these studies is the existence of between-subject variability in tau PET binding that can roughly be encapsulated into subtypes of prominent cortical tau, prominent medial-temporal tau, and typical AD.
In our view, the great contribution from Vogel and colleagues is the expansion of the topic in several directions. Firstly, they applied a data-driven method called SuStaIn. In this regard, it is important to enrich the current literature to assess the convergence between different subtyping methods, whether they are data-driven (Whitwell et al., 2018; Jeon et al., 2019; Vogel et al., 2021) or hypothesis-driven (Charil et al., 2019; Ossenkopelle et al., 2020; Mohanty et al., 2020), since the agreement between subtyping methods is currently low (Mohanty et al., 2020). Secondly, the authors assessed the largest cohort to date in a study of this kind, including 443 tau-PET-positive individuals, and a validation using an independent cohort of 469 individuals with a second tau tracer. Thirdly, the SuStaIn method seems to be the closest we can get today to understand the trajectories of tau spreading.
Should we now forget typical Alzheimer’s? In our view, the study by Vogel and colleagues does not suggest so. The authors nicely show between-subject variability in tau PET binding within a cohort where typical AD is the most common pattern of tau distribution. In our opinion, except for the MTL-sparing AD subtype (prevalence, 17 percent), all other subtypes in the study represent variations within typical AD. We agree that full-blown typical AD does not seem to emerge in earlier SuStaIn stages. This may be due to the large proportion of cognitively unimpaired subjects in their cohort. Nonetheless, all four subtypes do converge into typical AD at advanced SuStaIn stages.
A remaining question is, what are the underlying mechanisms for the trajectories of these subtypes? Vogel et al. get us closer to an answer, but the mystery cannot be solved using a cross-sectional method such as SuStaIn. To date, only one method for longitudinal clustering has been developed, indicating that many subjects converge into typical AD at advanced disease stages (Poulakis et al., 2021, in print). Longitudinal clustering approaches overcome the assumption that subjects remain in the same subtype when the disease advances, which is unrealistic but is supported by SuStaIn (Yang et al., 2021, in press).
We find it important to underline the recent recommendation by the IWG group that AD diagnosis is restricted to subjects with both positive AD biomarkers and a specific AD phenotype (Dubois et al., 2021). The current findings should thus be put into that context.
References:
Charil A, Shcherbinin S, Southekal S, Devous MD, Mintun M, Murray ME, Miller BB, Schwarz AJ. Tau Subtypes of Alzheimer's Disease Determined in vivo Using Flortaucipir PET Imaging. J Alzheimers Dis. 2019;71(3):1037-1048. PubMed.
Dubois B, Villain N, Frisoni GB, Rabinovici GD, Sabbagh M, Cappa S, Bejanin A, Bombois S, Epelbaum S, Teichmann M, Habert MO, Nordberg A, Blennow K, Galasko D, Stern Y, Rowe CC, Salloway S, Schneider LS, Cummings JL, Feldman HH. Clinical diagnosis of Alzheimer's disease: recommendations of the International Working Group. Lancet Neurol. 2021 Jun;20(6):484-496. Epub 2021 Apr 29 PubMed.
Ferreira D, Nordberg A, Westman E. Biological subtypes of Alzheimer disease: A systematic review and meta-analysis. Neurology. 2020 Mar 10;94(10):436-448. Epub 2020 Feb 11 PubMed.
Jeon S, Kang JM, Seo S, Jeong HJ, Funck T, Lee SY, Park KH, Lee YB, Yeon BK, Ido T, Okamura N, Evans AC, Na DL, Noh Y. Topographical Heterogeneity of Alzheimer's Disease Based on MR Imaging, Tau PET, and Amyloid PET. Front Aging Neurosci. 2019;11:211. Epub 2019 Aug 20 PubMed.
Mohanty R, Mårtensson G, Poulakis K, Muehlboeck JS, Rodriguez-Vieitez E, Chiotis K, Grothe MJ, Nordberg A, Ferreira D, Westman E. Comparison of subtyping methods for neuroimaging studies in Alzheimer's disease: a call for harmonization. Brain Commun. 2020;2(2):fcaa192. Epub 2020 Nov 9 PubMed.
Ossenkoppele R, Lyoo CH, Sudre CH, van Westen D, Cho H, Ryu YH, Choi JY, Smith R, Strandberg O, Palmqvist S, Westman E, Tsai R, Kramer J, Boxer AL, Gorno-Tempini ML, La Joie R, Miller BL, Rabinovici GD, Hansson O. Distinct tau PET patterns in atrophy-defined subtypes of Alzheimer's disease. Alzheimers Dement. 2020 Feb;16(2):335-344. Epub 2020 Jan 4 PubMed.
Vogel JW, Young AL, Oxtoby NP, Smith R, Ossenkoppele R, Strandberg OT, La Joie R, Aksman LM, Grothe MJ, Iturria-Medina Y, Alzheimer’s Disease Neuroimaging Initiative, Pontecorvo MJ, Devous MD, Rabinovici GD, Alexander DC, Lyoo CH, Evans AC, Hansson O. Four distinct trajectories of tau deposition identified in Alzheimer's disease. Nat Med. 2021 May;27(5):871-881. Epub 2021 Apr 29 PubMed.
Whitwell JL, Graff-Radford J, Tosakulwong N, Weigand SD, Machulda M, Senjem ML, Schwarz CG, Spychalla AJ, Jones DT, Drubach DA, Knopman DS, Boeve BF, Ertekin-Taner N, Petersen RC, Lowe VJ, Jack CR Jr, Josephs KA. [18 F]AV-1451 clustering of entorhinal and cortical uptake in Alzheimer's disease. Ann Neurol. 2018 Feb;83(2):248-257. Epub 2018 Feb 6 PubMed.
Ludwig Maximilians University
Ludwig Maximilian University
This is an impressive paper led by Jake Vogel and Oskar Hansson’s group, clearly demonstrating that the spatial patterns of tau pathology are more heterogeneous than commonly thought. In the to-date largest tau-PET sample of 1,612 individuals, they show four relatively common subtypes of tau pathology distribution (i.e., limbic-predominant, MTL sparing, posterior and lateral temporal), thereby questioning the concept of “typical” Alzheimer’s disease. Importantly, these subtypes are not only characterized by heterogeneous tau pathology distribution but also by different clinical characteristics, cognitive profiles, and progression rates, together supporting the view that the spatial distribution and spread of tau pathology is a key driver of clinical AD phenotypes.
The main finding of the paper matches well with previous postmortem and tau-PET studies, showing that tau pathology patterns can significantly deviate from the typical Braak-staging scheme, as described for early onset AD or rare, clinical AD variants, including logopenic primary progressive aphasia, posterior cortical atrophy, or dysexecutive AD (Murray et al., 2011; Ossenkoppele et al., 2016; Scholl et al., 2017; Sintini et al., 2019; Townley et al., 2020). Similarly, we and others have previously reported considerable heterogeneity in tau-PET patterns in the ADNI sample, with subtype-specific tau spreading patterns (Jones et al., 2017; Franzmeier et al., 2020).
While atypical tau distributions previously have been assumed to be relatively rare, the large sample used in the current study clearly illustrates that that tau subtypes are, in fact, relatively common, i.e., between 18-33 percent per subtype, and that rare clinical AD variants (e.g. PCA) may just represent “extreme” variants of these common tau subtypes. This view aligns well with the framework of AD as a spectrum disorder in which primary AD pathology can distribute along different neuronal networks and thus induce different types of cognitive symptoms and phenotypes (see also Jones et al., 2020).
The current findings are clinically important, because the heterogeneity in tau pathology may help clinical trials to determine patient-tailored clinical and neuroimaging endpoints, which may increase their sensitivity to detect treatment effects, as suggested by us previously (Franzmeier et al., 2020). The heuristic value of tau-PET subtyping for diagnostic and prognostic purposes will depend on the extent of within-subtype variability compared to between-subtype variability of tau pathology and cognitive profiles. The current work, however, means a big step toward individualized prediction of disease trajectories.
Furthermore, the work will be important as a starting point for future studies to investigate the origin of different tau trajectories, including developmental predisposition (Rogalski et al., 2008; Miller et al., 2019), or regional vulnerability (Grothe et al., 2018) and their resulting AD phenotypes.
References:
Franzmeier N, Dewenter A, Frontzkowski L, Dichgans M, Rubinski A, Neitzel J, Smith R, Strandberg O, Ossenkoppele R, Buerger K, Duering M, Hansson O, Ewers M. Patient-centered connectivity-based prediction of tau pathology spread in Alzheimer's disease. Sci Adv. 2020 Nov;6(48) Print 2020 Nov PubMed.
Grothe MJ, Sepulcre J, Gonzalez-Escamilla G, Jelistratova I, Schöll M, Hansson O, Teipel SJ, Alzheimer’s Disease Neuroimaging Initiative. Molecular properties underlying regional vulnerability to Alzheimer's disease pathology. Brain. 2018 Sep 1;141(9):2755-2771. PubMed.
Jones DT, Graff-Radford J, Lowe VJ, Wiste HJ, Gunter JL, Senjem ML, Botha H, Kantarci K, Boeve BF, Knopman DS, Petersen RC, Jack CR Jr. Tau, amyloid, and cascading network failure across the Alzheimer's disease spectrum. Cortex. 2017 Dec;97:143-159. Epub 2017 Oct 3 PubMed.
Miller ZA, Spina S, Pakvasa M, Rosenberg L, Watson C, Mandelli ML, Paredes MF, Joie R, Rabinovici GD, Rosen HJ, Grinberg LT, Huang EJ, Miller BL, Seeley WW, Gorno-Tempini ML. Cortical developmental abnormalities in logopenic variant primary progressive aphasia with dyslexia. Brain Commun. 2019;1(1):fcz027. Epub 2019 Oct 23 PubMed.
Murray ME, Graff-Radford NR, Ross OA, Petersen RC, Duara R, Dickson DW. Neuropathologically defined subtypes of Alzheimer's disease with distinct clinical characteristics: a retrospective study. Lancet Neurol. 2011 Sep;10(9):785-96. PubMed.
Ossenkoppele R, Schonhaut DR, Schöll M, Lockhart SN, Ayakta N, Baker SL, O'Neil JP, Janabi M, Lazaris A, Cantwell A, Vogel J, Santos M, Miller ZA, Bettcher BM, Vossel KA, Kramer JH, Gorno-Tempini ML, Miller BL, Jagust WJ, Rabinovici GD. Tau PET patterns mirror clinical and neuroanatomical variability in Alzheimer's disease. Brain. 2016 May;139(Pt 5):1551-67. Epub 2016 Mar 8 PubMed.
Jones D, Lowe V, Graff-Radford J, Botha H, Wiepert D, Murphy MC, Murray M, Senjem M, Gunter J, Wiste H, Boeve B, Knopman D, Petersen R, Jack C. Patterns of neurodegeneration in dementia reflect a global functional state space. medRxiv. November 12, 2020. medRxiv.
Rogalski E, Johnson N, Weintraub S, Mesulam M. Increased frequency of learning disability in patients with primary progressive aphasia and their first-degree relatives. Arch Neurol. 2008 Feb;65(2):244-8. PubMed.
Schöll M, Ossenkoppele R, Strandberg O, Palmqvist S, Swedish BioFINDER study, Jögi J, Ohlsson T, Smith R, Hansson O. Distinct 18F-AV-1451 tau PET retention patterns in early- and late-onset Alzheimer's disease. Brain. 2017 Sep 1;140(9):2286-2294. PubMed.
Sintini I, Martin PR, Graff-Radford J, Senjem ML, Schwarz CG, Machulda MM, Spychalla AJ, Drubach DA, Knopman DS, Petersen RC, Lowe VJ, Jack CR Jr, Josephs KA, Whitwell JL. Longitudinal tau-PET uptake and atrophy in atypical Alzheimer's disease. Neuroimage Clin. 2019;23:101823. Epub 2019 Apr 10 PubMed.
Townley RA, Graff-Radford J, Mantyh WG, Botha H, Polsinelli AJ, Przybelski SA, Machulda MM, Makhlouf AT, Senjem ML, Murray ME, Reichard RR, Savica R, Boeve BF, Drubach DA, Josephs KA, Knopman DS, Lowe VJ, Jack CR Jr, Petersen RC, Jones DT. Progressive dysexecutive syndrome due to Alzheimer's disease: a description of 55 cases and comparison to other phenotypes. Brain Commun. 2020;2(1):fcaa068. Epub 2020 May 27 PubMed.
University of Pittsburgh
In this important and well-powered tau PET study, Vogel, Hansson, and colleagues discovered four different patterns of tau deposition using machine learning. That individuals with these patterns had different cognitive profiles found in clinical practice supports the clinical relevance of their findings. These phenomenological findings support previous studies showing heterogeneity in the regions where tau deposition predominates in individuals, and raise important aspects to be addressed in subsequent studies, including those related to mechanisms of disease progression (e.g., what is leading to these different patterns of deposition (mechanically), and clinical trial designs. Trials using flortaucipir (FTP) PET uptake in regions of interest as a surrogate should take into account the different tau trajectories that individuals may follow, to try to minimize inter-individual variabilities in the regional rates of tau deposition. A modest rate of tau accumulation in a composite ROI could easily be confused with a therapeutic effect, if the individual's pattern of tau accumulation is not well captured by the chosen ROI).
It should be noted that it is well known that different tau PET tracers show different patterns of deposition, due to differences in sensitivity to tangles and off-target binding. FTP has repeatedly shown patterns of tau accumulation not very compatible with Braak stages. On the other hand, other tracers may show patterns of accumulation more similar to the patterns proposed by Braak and colleagues. Since factors such as off-target binding cannot be excluded from the results, it would be desirable to replicate their findings using different techniques. The partial replication of their results using RO948 greatly supported their findings, but the fact that RO948 subtypes presented some differences from FTP subtypes emphasizes the importance of future studies designed to refine the “subtypes” proposed.
This study is certainly a very important contribution to the field and paves the way for subsequent studies designed to replicate and refine these very important findings.
Katholieke Universiteit Leuven, Department of Imaging and Pathology, Laboratory of Neuropathology
KU Leuven
KU Leuven
The article by Vogel and colleagues indicates the existence of four spatiotemporal patterns in Alzheimer’s disease (AD), with a prevalence ranging between 18-33 percent each. The elegance of the SuStaIn approach applied here lies in the fact that these findings are entirely data-driven and not a priori defined by clinical diagnosis. Replication in an independent cohort, using a different PET ligand, reinforced the robustness of the study findings.
Compared to the seminal neuropathological study on AD subtypes (Murray et al., 2011), this in vivo, data-driven classification confirms in principle the three subtypes described by Murray et al. and splits the “typical AD” group into a posterior variant as well as a lateral temporal variant of AD. These subtypes align well with the most common atypical clinical presentations of AD, i.e., posterior cortical atrophy and logopenic variant of primary progressive aphasia (LV PPA), respectively. In the clinic, the presence of positive amyloid biomarkers (PET or CSF) in atypical AD variants has long been the only indication to prescribe cholinergic medication for a given patient (Mesulam et al., 2019; Schaeverbeke et al., 2017). Thus, the findings of Vogel et al. will help to improve the clinical diagnosis of AD and its subtypes, and to establish more individualized treatment strategies.
However, co-pathologies should not be forgotten. In addition to tau-based subclassifications, neuropathologists described other pathologies that co-exist in AD, specifically cerebral amyloid angiopathy (CAA), α-synuclein, and TDP-43 pathology (more recently described as LATE-NC) (Josephs et al., 2016; Thal et al., 2003; Toledo et al., 2016). Here, interactions of these co-pathologies with amyloid-β and/or tau pathology have been reported, including interactions with the prion protein (Gomes et al., 2019; Spires-Jones et al., 2017; Tomé et al., 2021). This suggests that these pathologies contribute to the clinical manifestation of dementia in the elderly and probably influence the tau-PET tracer binding pattern. Capillary CAA has previously been identified to indicate a distinct type of AD strongly associated with the presence of the APOE ε4 allele (Thal et al., 2010), which might correspond to the limbic type shown by Vogel et al., given that this subtype was also associated with APOE ε4. On top of that, by focusing on lesion morphology (Josephs et al., 2019) and TDP-43 phosphorylation patterns (Tomé et al., 2020), distinct TDP-43 neuropathological patterns have been identified in AD cases.
Questions then arise as to whether and how these co-morbid pathologies, and their distributions, are associated with the four subtypes identified by Vogel and colleagues. Is a prediction possible for the underlying neuropathological picture when going beyond tau?
And still, a caveat to all this needs to be taken into account: Tau PET detects only a small part of the tau pathology observed neuropathologically, which is usually restricted to Braak neurofibrillary tangle (NFT) stage V and VI cases (Soleimani-Meigooni et al., 2020). Therefore, the patterns reported by Vogel et al. are restricted to cases that may already be present with a (nearly) full-blown neuropathological picture of tau pathology related to the Braak NFT stage. However, the neuropathological subtypes defined by Murray et al. do not vary among Braak NFT stages. This indicates, in our opinion, that these AD subtypes do not challenge the neuropathological staging of tau pathology but add specific subtypes on top, which can be diagnosed mainly in an advanced stage of the underlying tau pathology. When focusing on prevention strategies that require a diagnosis of preclinical AD, the tau-PET based subtyping therefore may be of less help.
Overall, this study may have an important impact on stratifying symptomatic AD patients for clinical trials in order to find the proper subgroup of patients eligible for a given treatment strategy. However, preclinical stages may be missed from these subtyping strategies. Future work will need to focus on the contribution of co-pathologies in these AD subtypes to allow a proper stratification in clinical trials.
References:
Gomes LA, Hipp SA, Rijal Upadhaya A, Balakrishnan K, Ospitalieri S, Koper MJ, Largo-Barrientos P, Uytterhoeven V, Reichwald J, Rabe S, Vandenberghe R, von Arnim CA, Tousseyn T, Feederle R, Giudici C, Willem M, Staufenbiel M, Thal DR. Aβ-induced acceleration of Alzheimer-related τ-pathology spreading and its association with prion protein. Acta Neuropathol. 2019 Dec;138(6):913-941. Epub 2019 Aug 14 PubMed.
Josephs KA, Murray ME, Tosakulwong N, Weigand SD, Serie AM, Perkerson RB, Matchett BJ, Jack CR Jr, Knopman DS, Petersen RC, Parisi JE, Petrucelli L, Baker M, Rademakers R, Whitwell JL, Dickson DW. Pathological, imaging and genetic characteristics support the existence of distinct TDP-43 types in non-FTLD brains. Acta Neuropathol. 2019 Feb;137(2):227-238. Epub 2019 Jan 2 PubMed.
Josephs KA, Murray ME, Whitwell JL, Tosakulwong N, Weigand SD, Petrucelli L, Liesinger AM, Petersen RC, Parisi JE, Dickson DW. Updated TDP-43 in Alzheimer's disease staging scheme. Acta Neuropathol. 2016 Apr;131(4):571-85. Epub 2016 Jan 25 PubMed.
Mesulam MM, Lalehzari N, Rahmani F, Ohm D, Shahidehpour R, Kim G, Gefen T, Weintraub S, Bigio E, Geula C. Cortical cholinergic denervation in primary progressive aphasia with Alzheimer pathology. Neurology. 2019 Apr 2;92(14):e1580-e1588. Epub 2019 Mar 6 PubMed.
Murray ME, Graff-Radford NR, Ross OA, Petersen RC, Duara R, Dickson DW. Neuropathologically defined subtypes of Alzheimer's disease with distinct clinical characteristics: a retrospective study. Lancet Neurol. 2011 Sep;10(9):785-96. PubMed.
Nelson PT, Dickson DW, Trojanowski JQ, Jack CR, Boyle PA, Arfanakis K, Rademakers R, Alafuzoff I, Attems J, Brayne C, Coyle-Gilchrist IT, Chui HC, Fardo DW, Flanagan ME, Halliday G, Hokkanen SR, Hunter S, Jicha GA, Katsumata Y, Kawas CH, Keene CD, Kovacs GG, Kukull WA, Levey AI, Makkinejad N, Montine TJ, Murayama S, Murray ME, Nag S, Rissman RA, Seeley WW, Sperling RA, White Iii CL, Yu L, Schneider JA. Limbic-predominant age-related TDP-43 encephalopathy (LATE): consensus working group report. Brain. 2019 Jun 1;142(6):1503-1527. PubMed.
Schaeverbeke J, Evenepoel C, Bruffaerts R, Van Laere K, Bormans G, Dries E, Tousseyn T, Nelissen N, Peeters R, Vandenbulcke M, Dupont P, Vandenberghe R. Cholinergic depletion and basal forebrain volume in primary progressive aphasia. Neuroimage Clin. 2017;13:271-279. Epub 2016 Dec 2 PubMed.
Soleimani-Meigooni DN, Iaccarino L, La Joie R, Baker S, Bourakova V, Boxer AL, Edwards L, Eser R, Gorno-Tempini ML, Jagust WJ, Janabi M, Kramer JH, Lesman-Segev OH, Mellinger T, Miller BL, Pham J, Rosen HJ, Spina S, Seeley WW, Strom A, Grinberg LT, Rabinovici GD. 18F-flortaucipir PET to autopsy comparisons in Alzheimer's disease and other neurodegenerative diseases. Brain. 2020 Dec 5;143(11):3477-3494. PubMed.
Spires-Jones TL, Attems J, Thal DR. Interactions of pathological proteins in neurodegenerative diseases. Acta Neuropathol. 2017 Aug;134(2):187-205. Epub 2017 Apr 11 PubMed.
Thal DR, Ghebremedhin E, Orantes M, Wiestler OD. Vascular pathology in Alzheimer disease: correlation of cerebral amyloid angiopathy and arteriosclerosis/lipohyalinosis with cognitive decline. J Neuropathol Exp Neurol. 2003 Dec;62(12):1287-301. PubMed.
Thal DR, Papassotiropoulos A, Saido TC, Griffin WS, Mrak RE, Kölsch H, Del Tredici K, Attems J, Ghebremedhin E. Capillary cerebral amyloid angiopathy identifies a distinct APOE epsilon4-associated subtype of sporadic Alzheimer's disease. Acta Neuropathol. 2010 Aug;120(2):169-83. PubMed.
Toledo JB, Gopal P, Raible K, Irwin DJ, Brettschneider J, Sedor S, Waits K, Boluda S, Grossman M, Van Deerlin VM, Lee EB, Arnold SE, Duda JE, Hurtig H, Lee VM, Adler CH, Beach TG, Trojanowski JQ. Pathological α-synuclein distribution in subjects with coincident Alzheimer's and Lewy body pathology. Acta Neuropathol. 2016 Mar;131(3):393-409. Epub 2015 Dec 31 PubMed.
Tomé SO, Gomes LA, Li X, Vandenberghe R, Tousseyn T, Thal DR. TDP-43 interacts with pathological τ protein in Alzheimer's disease. Acta Neuropathol. 2021 May;141(5):795-799. Epub 2021 Apr 2 PubMed.
Tomé SO, Vandenberghe R, Ospitalieri S, Van Schoor E, Tousseyn T, Otto M, von Arnim CA, Thal DR. Distinct molecular patterns of TDP-43 pathology in Alzheimer's disease: relationship with clinical phenotypes. Acta Neuropathol Commun. 2020 Apr 29;8(1):61. PubMed.
Make a Comment
To make a comment you must login or register.