Does Astrocyte Tau Cause Dementia?
Quick Links
Once again, tau creates havoc—this time by hijacking astrocytes. According to a November 9 report in Nature Neuroscience, researchers led by Nicolas Toni at the University of Lausanne, Switzerland, found that in people with AD, tau accumulated in astrocytes of the hilus, a portion of the hippocampus. In mice, they found that too much tau in these astrocytes impaired mitochondrial function and ATP production. This reduced the number of inhibitory neurons in the hippocampus, put the kibosh on neurogenesis, and suppressed neuronal network activity there. The mice had trouble navigating a water maze. Mouse memory deficits were rescued by ramping up activity of inhibitory neurons. All told, the findings hint that tau pathology extends beyond neurons in people with AD and that glial tau might contribute to dementia.
- In AD, 3R tau accumulates in hippocampal astrocytes.
- In mouse hippocampal astrocytes, 3R tau damages mitochondria.
- This weakens neuronal network activity and spatial memory.
Alberto Serrano-Pozo and Bradley Hyman of Massachusetts General Hospital, Charlestown, were intrigued by these findings but wondered about the origin of the tau. “Is tau expression increased in AD reactive astrocytes, or is it taken up from the extracellular space after being released by neurons?” they wondered (see full comment below). This remains to be determined, though Richetin noted that they only saw astrocytes with three-repeat (3R) tau in hippocampal regions that also had neurofibrillary tangles, hinting that the protein may have come from the neurons.
Tau tangles are an integral part of AD pathology, appearing in the hippocampus in early stage disease and then gradually spreading throughout the brain. Their accumulation closely mirrors cognitive decline (Braak and Braak, 1991; Jun 2019 news; May 2019 news). Researchers have focused on tau’s role in neuron dysfunction and death (Jan 2020 news; Aug 2016 news). What about other cells?
First author Kevin Richetin looked for tau tangles in different areas of the hippocampus in tissue samples from healthy controls and from people with AD. As expected, he found them in the AD samples, but the pathology was not equally distributed; there were hot spots in certain areas, including the hilus.
“The hilus is seen as a highway between the dentate gyrus and CA3 region within the hippocampus, so most researchers do not pay much attention to what happens there,” Toni told Alzforum. “It turns out to be very important.”
Zooming in on the astrocytes in the hilus, Richetin saw they were packed with 3R tau in the AD tissue samples. This isoform contributes to the 3R/4R type of neurofibrillary tangles found in Alzheimer’s. The amount of 3R tau in the astrocytes correlated with tau tangles and with Aβ plaques in the surrounding tissue (see image below), suggesting the 3R tau accumulation may be downstream of other AD pathologies. Oddly enough, Richetin found no increase in phosphorylated forms of tau in the astrocytes or any evidence of tangles in these cells. He speculated that this might be because these hilar astrocytes cells do not phosphorylate the protein as easily as neurons do, or that tau is dephosphorylated by the astrocytes.

Tau-Stuffed Astrocytes. Within the hippocampus, 3R tau (red) in astrocytes correlated with amyloid plaque burden and levels of phospho-tau181. [Courtesy of Richetin et al., Nature Neuroscience, 2020.]
Understanding Astrocytic Tau Accumulations
Phosphorylated tau and tangles have been found in astrocytes in disorders such as age-related tau astrogliopathy (ARTAG), progressive supranuclear palsy (PSP), and corticobasal degeneration (CBD) (Kovacs et al., 2016; Briel et al., 2020). However, this astrocytic tau is poorly understood, especially in AD.
Does this 3R tau accumulation affect the astrocytes or their surrounding neurons? To test this idea in a mouse model, Richetin and colleagues collaborated with Nicole Déglon, also at the University of Lausanne, to overproduce tau in mouse hippocampi. Previously, Déglon worked out how to selectively express genes in astrocytes using lentivirus vectors, and the researchers used this same strategy for tau (Humbel et al., 2020). Richetin injected lentiviruses carrying a gene for human 3R tau into the hippocampi of 3-month-old wild-type mice. Two weeks later, he verified that the human 3R tau was expressed only in hilar astrocytes.
Four months after the lentiviral injection, Richetin found that the hilar astrocytes were packed with 3R tau. Richetin found that within these astrocytes, mitochondria languished in the cell bodies, rather than travelling to the astrocyte arms that support neurons. Mitochondrial function also suffered; the organelles replenished less often, and they produced much less ATP than usual. “If there are no dynamic mitochondria in astrocytes, they have no power to support brain activity,” Richetin noted.
Still, neurons looked normal for the most part, with no signs of neuronal death. However, neurogenesis had faltered, and the treated mice had fewer parvalbumin-positive inhibitory neurons in the dentate gyrus than controls. The number of inhibitory synapses also collapsed.
Parvalbumin-producing neurons are like the pacemakers of the brain, modulating γ-frequency oscillations, which is important for working memory (Kim et al., 2015). Indeed, the 3R mice had weaker γ activity in the hippocampus and Richetin found they had trouble finding a hidden platform in a water maze. Otherwise, they seemed to behave normally.
Taken together, the results suggested that accumulation of tau in hilar astrocytes compromised the function of hippocampal inhibitory neurons. “Neurons and astrocytes are so codependent and intermingled that it is hard to tease them apart,” Toni noted. “To know that the phenotype we observed is only due to tau accumulation in astrocytes, not neurons, is critically important.”
The researchers were also able to correct the spatial-memory deficit by injecting the neurotrophin neuregulin 1 (NRG1) into the hippocampus. This synaptic cleft protein stimulated production of parvalbumin by inhibitory neurons and improved spatial navigation in the 3R mice (see image below). NRG1 is known to increase the activity of parvalbumin-positive inhibitory neurons (Li et al., 2011).
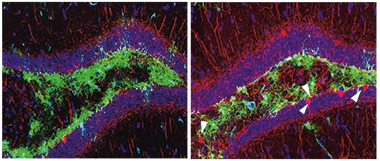
Restoring Par. In the hippocampi of 3R tau mice (left), parvalbumin-positive inhibitory neurons (red) are sparse and memory falters. Injecting the hippocampus with neuregulin 1 ramps up parvalbumin production (right) and rescues spatial navigation (not shown). [Courtesy of Richetin et al., Nature Neuroscience, 2020.]
Toni was surprised that altering just a few hundred astrocytes could have such a profound effect in mice. Whether that might hold true in people remains to be seen. “The human hippocampus is about as big as the pinky finger and the hilus is about one-tenth of that, with one-third of its cells being astrocytes. If we could target this minute number of cells and have such a profound effect on behavior, it would be pretty amazing,” he said.
In fact, researchers at MIT led by Li-Huei Tsai are testing γ-frequency strengthening strategies in clinical trials. Tsai previously reported that this preserves cortical neurons and stimulates microglia to gobble up amyloid plaques (May 2019 news). Tsai found this new study intriguing but said it raises many questions. She wondered if microglia also accumulate 3R tau, and if the astrocyte tau trove was limited to the 3R isoform.—Chelsea Weidman Burke
References
News Citations
- Serial PET Nails It: Preclinical AD Means Amyloid, Tau, then Cognitive Decline
- Longitudinal Tau PET Links Aβ to Subsequent Rise in Cortical Tau
- Does Tau Kill Neurons by Way of Necroptosis?
- No Special Glasses Needed: Three-Dimensional Views of Tau and Aβ in the Brain
- Gamma Waves Synchronized by Light: Good for Synapses, Memory?
Paper Citations
- Braak H, Braak E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991;82(4):239-59. PubMed.
- Kovacs GG, Ferrer I, Grinberg LT, Alafuzoff I, Attems J, Budka H, Cairns NJ, Crary JF, Duyckaerts C, Ghetti B, Halliday GM, Ironside JW, Love S, Mackenzie IR, Munoz DG, Murray ME, Nelson PT, Takahashi H, Trojanowski JQ, Ansorge O, Arzberger T, Baborie A, Beach TG, Bieniek KF, Bigio EH, Bodi I, Dugger BN, Feany M, Gelpi E, Gentleman SM, Giaccone G, Hatanpaa KJ, Heale R, Hof PR, Hofer M, Hortobágyi T, Jellinger K, Jicha GA, Ince P, Kofler J, Kövari E, Kril JJ, Mann DM, Matej R, McKee AC, McLean C, Milenkovic I, Montine TJ, Murayama S, Lee EB, Rahimi J, Rodriguez RD, Rozemüller A, Schneider JA, Schultz C, Seeley W, Seilhean D, Smith C, Tagliavini F, Takao M, Thal DR, Toledo JB, Tolnay M, Troncoso JC, Vinters HV, Weis S, Wharton SB, White CL 3rd, Wisniewski T, Woulfe JM, Yamada M, Dickson DW. Aging-related tau astrogliopathy (ARTAG): harmonized evaluation strategy. Acta Neuropathol. 2016 Jan;131(1):87-102. Epub 2015 Dec 10 PubMed.
- Briel N, Pratsch K, Roeber S, Arzberger T, Herms J. Contribution of the astrocytic tau pathology to synapse loss in progressive supranuclear palsy and corticobasal degeneration. Brain Pathol. 2020 Oct 22;:e12914. PubMed.
- Humbel M, Ramosaj M, Zimmer V, Regio S, Aeby L, Moser S, Boizot A, Sipion M, Rey M, Déglon N. Maximizing lentiviral vector gene transfer in the CNS. Gene Ther. 2020 Jul 6; PubMed.
- Kim T, Thankachan S, McKenna JT, McNally JM, Yang C, Choi JH, Chen L, Kocsis B, Deisseroth K, Strecker RE, Basheer R, Brown RE, McCarley RW. Cortically projecting basal forebrain parvalbumin neurons regulate cortical gamma band oscillations. Proc Natl Acad Sci U S A. 2015 Mar 17;112(11):3535-40. Epub 2015 Mar 2 PubMed.
- Li KX, Lu YM, Xu ZH, Zhang J, Zhu JM, Zhang JM, Cao SX, Chen XJ, Chen Z, Luo JH, Duan S, Li XM. Neuregulin 1 regulates excitability of fast-spiking neurons through Kv1.1 and acts in epilepsy. Nat Neurosci. 2011 Dec 11;15(2):267-73. PubMed.
Further Reading
Papers
- Nolan A, De Paula Franca Resende E, Petersen C, Neylan K, Spina S, Huang E, Seeley W, Miller Z, Grinberg LT. Astrocytic Tau Deposition Is Frequent in Typical and Atypical Alzheimer Disease Presentations. J Neuropathol Exp Neurol. 2019 Dec 1;78(12):1112-1123. PubMed.
- Reid MJ, Beltran-Lobo P, Johnson L, Perez-Nievas BG, Noble W. Astrocytes in Tauopathies. Front Neurol. 2020;11:572850. Epub 2020 Sep 24 PubMed.
Primary Papers
- Richetin K, Steullet P, Pachoud M, Perbet R, Parietti E, Maheswaran M, Eddarkaoui S, Bégard S, Pythoud C, Rey M, Caillierez R, Q Do K, Halliez S, Bezzi P, Buée L, Leuba G, Colin M, Toni N, Déglon N. Tau accumulation in astrocytes of the dentate gyrus induces neuronal dysfunction and memory deficits in Alzheimer's disease. Nat Neurosci. 2020 Dec;23(12):1567-1579. Epub 2020 Nov 9 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Massachusetts General Hospital / Harvard Medical School
In this important study, Richetin et al. demonstrate that astrocytes in the hilus of the hippocampus dentate gyrus accumulate misfolded 3R-tau in Alzheimer’s disease (AD). Using an elegant lentiviral gene transfer approach in vivo (wild-type mice) and in vitro (rat hippocampal neuron/astrocyte co-cultures and mouse acute hippocampal slices), they studied the downstream effects of this tau accumulation in hilar astrocytes and observed multiple phenotypes in both astrocytes and neurons when overexpressing 1N3R-tau in astrocytes: impairment to astrocyte mitochondrial transport, to REDOX and ATP production, as well as to calcium homeostasis; reduced maturation of newborn neurons in the subgranular zone of the dentate gyrus; structural synaptic alterations involving excitatory and inhibitory synapses; loss of inhibitory parvalbumin-positive neurons, and impaired hippocampal circuit γ oscillations and spatial memory.…More
These findings highlight the astrocyte cell-autonomous and non-cell-autonomous consequences in the setting of neurodegenerative proteinopathies, and support the idea that the biology of reactive astrocytes, not just neurons, is negatively impacted by protein aggregates in AD and other tauopathies, leading to further neuronal dysfunction.
Some questions remain to be elucidated. Is tau expression increased in AD-reactive astrocytes or is it taken up from the extracellular space by astrocytes after being released by neurons? Astrocytes express very low levels of MAPT, encoding the microtubule-associated protein tau, compared to neurons in the normal brain (Zhang et al., 2016). Recent single-nuclei RNA-Seq studies have not reported increased levels of MAPT transcripts in AD-reactive astrocytes (Mathys et al., 2019; Grubman et al., 2019; Zhou et al., 2020). However, tau-immunoreactive astrocytes can be found in the brains of otherwise normal aged individuals and more frequently in subjects with AD and other neurodegenerative diseases, with granular fuzzy and thorn-shaped morphologies, and within subpial, perivascular, subependymal, white-matter and gray-matter areas, including the medial temporal lobe (Lace et al., 2012; López-González et al., 2012)—collectively called Aging-Related Tau Astrogliopathy, or ARTAG (Kovacs et al., 2017).
In addition, tau-immunoreactive astrocytes are pathological hallmarks of the primary tauopathies progressive supranuclear palsy and corticobasal degeneration, which exhibit tufted astrocytes and astrocytic plaques, respectively (Dickson et al., 1999), and they are also conspicuous in chronic traumatic encephalopathy (CTE) (McKee et al., 2016). Intriguingly, the astrocytic tau inclusions described in all these conditions contain primarily 4R-tau rather than 3R-tau. It is unclear why 3R-tau staining was more prominent than 4R-tau in the hilar astrocytes of this study's subjects, and why the accumulation of 3R-tau was more deleterious than that of 4R-tau for both astrocytes and neurons.
What is the mechanism of tau uptake by astrocytes? Neurofibrillary tangles develop primarily in glutamatergic pyramidal neurons and misfolded tau is thought to propagate through neural circuits trans-synaptically. Astrocyte fine processes rich in glutamate transporters are a structural and functional part of the excitatory glutamatergic synapses (Ventura and Harris, 1999). Thus, it is plausible that astrocytes, which are phagocytic cells, take up misfolded tau released by neurons at excitatory synapses. In addition, reactive astrocytes actively infiltrate extracellular “ghost” tangles with their processes (Serrano-Pozo et al., 2011; Irwin et al., 2012; Perez-Nievas and Serrano-Pozo, 2018), and can phagocytose amyloid-plaque-associated neuritic dystrophies (Gomez-Arboledas et al., 2018), which contain tau aggregates. Recently, the low-density lipoprotein related protein 1 (LRP1) has been involved in tau uptake by neurons (Rauch et al., 2020), but LRP1 is also highly expressed in astrocytes among other cell types, and upregulated in AD-reactive astrocytes (Arélin et al., 2002). Indeed, knocking down LRP1 specifically in neurons reduced trans-synaptic tau propagation through neural circuit but increased tau uptake by astrocytes (Rauch et al., 2020).
In summary, Richetin et al.’s data show that there are profound synergies between astrocytes and neurons in the pathophysiology of AD and other tauopathies, and suggest that a deeper understanding of astrocytic alterations may prove to be critical in understanding neurodegeneration.
References:
Zhang Y, Sloan SA, Clarke LE, Caneda C, Plaza CA, Blumenthal PD, Vogel H, Steinberg GK, Edwards MS, Li G, Duncan JA 3rd, Cheshier SH, Shuer LM, Chang EF, Grant GA, Gephart MG, Barres BA. Purification and Characterization of Progenitor and Mature Human Astrocytes Reveals Transcriptional and Functional Differences with Mouse. Neuron. 2016 Jan 6;89(1):37-53. Epub 2015 Dec 10 PubMed.
Mathys H, Davila-Velderrain J, Peng Z, Gao F, Mohammadi S, Young JZ, Menon M, He L, Abdurrob F, Jiang X, Martorell AJ, Ransohoff RM, Hafler BP, Bennett DA, Kellis M, Tsai LH. Author Correction: Single-cell transcriptomic analysis of Alzheimer's disease. Nature. 2019 Jul;571(7763):E1. PubMed.
Grubman A, Chew G, Ouyang JF, Sun G, Choo XY, McLean C, Simmons RK, Buckberry S, Vargas-Landin DB, Poppe D, Pflueger J, Lister R, Rackham OJ, Petretto E, Polo JM. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nat Neurosci. 2019 Dec;22(12):2087-2097. PubMed.
Zhou Y, Song WM, Andhey PS, Swain A, Levy T, Miller KR, Poliani PL, Cominelli M, Grover S, Gilfillan S, Cella M, Ulland TK, Zaitsev K, Miyashita A, Ikeuchi T, Sainouchi M, Kakita A, Bennett DA, Schneider JA, Nichols MR, Beausoleil SA, Ulrich JD, Holtzman DM, Artyomov MN, Colonna M. Author Correction: Human and mouse single-nucleus transcriptomics reveal TREM2-dependent and TREM2-independent cellular responses in Alzheimer's disease. Nat Med. 2020 Jun;26(6):981. PubMed.
Lace G, Ince PG, Brayne C, Savva GM, Matthews FE, de Silva R, Simpson JE, Wharton SB. Mesial Temporal Astrocyte Tau Pathology in the MRC-CFAS Ageing Brain Cohort. Dement Geriatr Cogn Disord. 2012;34(1):15-24. PubMed.
López-González I, Carmona M, Blanco R, Luna-Muñoz J, Martínez-Mandonado A, Mena R, Ferrer I. Characterization of Thorn-Shaped Astrocytes in White Matter of Temporal Lobe in Alzheimer's Disease Brains. Brain Pathol. 2012 Aug 13; PubMed.
Kovacs GG, Robinson JL, Xie SX, Lee EB, Grossman M, Wolk DA, Irwin DJ, Weintraub D, Kim CF, Schuck T, Yousef A, Wagner ST, Suh E, Van Deerlin VM, Lee VM, Trojanowski JQ. Evaluating the Patterns of Aging-Related Tau Astrogliopathy Unravels Novel Insights Into Brain Aging and Neurodegenerative Diseases. J Neuropathol Exp Neurol. 2017 Apr 1;76(4):270-288. PubMed.
Dickson DW. Neuropathologic differentiation of progressive supranuclear palsy and corticobasal degeneration. J Neurol. 1999 Sep;246 Suppl 2:II6-15. PubMed.
McKee AC, Cairns NJ, Dickson DW, Folkerth RD, Keene CD, Litvan I, Perl DP, Stein TD, Vonsattel JP, Stewart W, Tripodis Y, Crary JF, Bieniek KF, Dams-O'Connor K, Alvarez VE, Gordon WA, TBI/CTE group. The first NINDS/NIBIB consensus meeting to define neuropathological criteria for the diagnosis of chronic traumatic encephalopathy. Acta Neuropathol. 2016 Jan;131(1):75-86. Epub 2015 Dec 14 PubMed.
Ventura R, Harris KM. Three-dimensional relationships between hippocampal synapses and astrocytes. J Neurosci. 1999 Aug 15;19(16):6897-906. PubMed.
Serrano-Pozo A, Mielke ML, Gómez-Isla T, Betensky RA, Growdon JH, Frosch MP, Hyman BT. Reactive glia not only associates with plaques but also parallels tangles in Alzheimer's disease. Am J Pathol. 2011 Sep;179(3):1373-84. PubMed.
Irwin DJ, Cohen TJ, Grossman M, Arnold SE, Xie SX, Lee VM, Trojanowski JQ. Acetylated tau, a novel pathological signature in Alzheimer's disease and other tauopathies. Brain. 2012 Mar;135(Pt 3):807-18. PubMed.
Perez-Nievas BG, Serrano-Pozo A. Deciphering the Astrocyte Reaction in Alzheimer's Disease. Front Aging Neurosci. 2018;10:114. Epub 2018 Apr 25 PubMed.
Gomez-Arboledas A, Davila JC, Sanchez-Mejias E, Navarro V, Nuñez-Diaz C, Sanchez-Varo R, Sanchez-Mico MV, Trujillo-Estrada L, Fernandez-Valenzuela JJ, Vizuete M, Comella JX, Galea E, Vitorica J, Gutierrez A. Phagocytic clearance of presynaptic dystrophies by reactive astrocytes in Alzheimer's disease. Glia. 2018 Mar;66(3):637-653. Epub 2017 Nov 27 PubMed.
Rauch JN, Luna G, Guzman E, Audouard M, Challis C, Sibih YE, Leshuk C, Hernandez I, Wegmann S, Hyman BT, Gradinaru V, Kampmann M, Kosik KS. LRP1 is a master regulator of tau uptake and spread. Nature. 2020 Apr;580(7803):381-385. Epub 2020 Apr 1 PubMed.
Arélin K, Kinoshita A, Whelan CM, Irizarry MC, Rebeck GW, Strickland DK, Hyman BT. LRP and senile plaques in Alzheimer's disease: colocalization with apolipoprotein E and with activated astrocytes. Brain Res Mol Brain Res. 2002 Jul 15;104(1):38-46. PubMed.
University of Kentucky
This is a very interesting study that highlights the susceptibility of astrocytes to preferentially acquire pathological features in the form of confirmation-specific isoforms of tau in hippocampal regions burdened by Alzheimer pathology.
Seeking to understand the functional consequence of astrocytes burdened by 3R tau, the authors demonstrate that viral-driven expression of the 3R isoform alters mitochondrial transport. Similar anterograde disruptions in the axonal movement of APP were observed in neuronal cultures when the balance of 3R:4R tau was manipulated (Lacovich et al., 2017).
Here, Richetin et al. demonstrate that driving astrocyte-specific expression of 3R in wild-type C57BL6/J mice is sufficient to drive dysfunctional responses of interneurons, manifesting as cognitive deficits. Given that the wild-type mouse expresses only the 4R confirmation in the adult brain, compared to equimolar distribution of 3R:4R in humans (Andreadis 2005), it would be interesting to know whether these approaches drove the same manifestations in mouse models with humanized MAPT.…More
Further, whether accumulation of 4R tau in astrocytes is sufficient to alter neuronal and cognitive dysfunction was not fully elaborated in the current work. Such studies would provide potential mechanistic or dysfunctional underpinnings of astrocytes that acquire predominantly 4R confirmations, as has been documented for ARTAG (Kovacs 2016).
Massachusetts General Hospital
This elegant study by Richetin and colleagues has several interesting and important findings.
In human participants with Alzheimer's disease, 3R tau accumulation was observed in astrocytes in the hippocampus, specifically the hilus of the dentate gyrus. The amount of 3R (but not 4R) tau in hilar astrocytes was positively correlated with Braak stage, amyloid burden, and synaptic alterations that were specific to the postsynaptic side (increased expression of PSD95).
Additionally, mouse models convincingly demonstrated that tau pathology in hilar astrocytes (induced via a novel lentiviral vector strategy) led to mitochondrial dysfunction, neurodegeneration, and spatial memory impairment.
This study builds on previous observations of AD-related tau accumulation in astrocytes by investigating the downstream consequences of this particular pathology, and adds to a growing body of evidence that glial cells are key players in the etiology of AD.…More
Sylics
This is a very intriguing and detailed analysis regarding the functional consequences of 3R tau in astrocytes. It is an important reminder that much remains to be learned about the functional consequences of pathological tau in astrocytes. Several older studies have pioneered the study of the consequences of astrocytic tau pathology using mouse models, but to our knowledge this work has never been followed up (Dabir et al., 2006; Forman et al., 2005).
We recently presented our findings regarding AAV-mediated viral delivery of truncated tau to astrocytes at AAIC Neuroscience Next 2020. In that study, we expressed human truncated tau (aa151-391/4R)—which we have previously shown to lead to neurofibrillary tangles in neurons (Vogels et al., 2020)—along with non-fused mCherry in a 1:1 ratio under the astrocytic GFAP promotor. In contrast to the study by Richetin and colleagues, we decided to focus on 4R tau, because astrocytic tau pathology in primary tauopathies is almost exclusively associated with accumulation of 4R tau (Kovacs, 2020). …More
At five months after injection in hippocampi of wild-type mice, we found widespread transduction of astrocytes in all subregions of the hippocampus and overlying cortex. We also observed accumulation of p-tau S214 and T231 in astrocytes, but no AT8+ or Methoxy-XO4+ inclusions. No detectable cognitive deficits were observed in open field, novel object recognition, and Y-maze spontaneous alterations tests. At five months after AAV injection in cortex, we also did not detect alterations in neuronal activity in the vicinity of truncated tau positive astrocytes (as visualized with mCherry co-expression) using in vivo 2-photon calcium imaging.
More studies are therefore required to elucidate the effects of different tau species on astrocytes, the origins of tau in astrocytes, and the multitude of potential functional consequences of astrocytic tau pathology, e.g. on the vasculature.
References:
Dabir DV, Robinson MB, Swanson E, Zhang B, Trojanowski JQ, Lee VM, Forman MS. Impaired glutamate transport in a mouse model of tau pathology in astrocytes. J Neurosci. 2006 Jan 11;26(2):644-54. PubMed.
Forman MS, Lal D, Zhang B, Dabir DV, Swanson E, Lee VM, Trojanowski JQ. Transgenic mouse model of tau pathology in astrocytes leading to nervous system degeneration. J Neurosci. 2005 Apr 6;25(14):3539-50. PubMed.
Kovacs GG. Astroglia and Tau: New Perspectives. Front Aging Neurosci. 2020;12:96. Epub 2020 Apr 9 PubMed.
Vogels T, Vargová G, Brezováková V, Quint WH, Hromádka T. Viral Delivery of Non-Mutated Human Truncated Tau to Neurons Recapitulates Key Features of Human Tauopathy in Wild-Type Mice. J Alzheimers Dis. 2020;77(2):551-568. PubMed.
A recent study in Nature finds that microglia sense ATP and then inhibit neuronal activity (Badimon et al., 2020). Thus, if there is less ATP production by astrocytes then maybe there is also less inhibition by microglia? Additionally, another new Nature paper finds that astrocytes control excitatory and inhibitory synapses (Takano et al., 2020).
References:
Badimon A, Strasburger HJ, Ayata P, Chen X, Nair A, Ikegami A, Hwang P, Chan AT, Graves SM, Uweru JO, Ledderose C, Kutlu MG, Wheeler MA, Kahan A, Ishikawa M, Wang YC, Loh YE, Jiang JX, Surmeier DJ, Robson SC, Junger WG, Sebra R, Calipari ES, Kenny PJ, Eyo UB, Colonna M, Quintana FJ, Wake H, Gradinaru V, Schaefer A. Negative feedback control of neuronal activity by microglia. Nature. 2020 Oct;586(7829):417-423. Epub 2020 Sep 30 PubMed. …More
Takano T, Wallace JT, Baldwin KT, Purkey AM, Uezu A, Courtland JL, Soderblom EJ, Shimogori T, Maness PF, Eroglu C, Soderling SH. Chemico-genetic discovery of astrocytic control of inhibition in vivo. Nature. 2020 Nov 11; PubMed.
Make a Comment
To make a comment you must login or register.