Most Detailed Look Yet at Activation States of Human Microglia
Quick Links
It is difficult to isolate large quantities of microglia from human brain. That’s why scientists still know little about the different ways these cells rear up in health and disease. A new single-cell—not single-nucleus—RNA-sequencing study led by Philip De Jager at Columbia University, New York, sheds some light on this. It analyzed the largest number of microglia yet, totaling more than 16,000, isolated from the brains of people with Alzheimer’s disease and normal cognition. In the November 30 Nature Communications, the authors describe nine distinct transcriptional states found in all brain samples. At least 80 percent of microglia fell into one of two homeostatic subtypes; that was true even in AD brains. Seven other subtypes comprised less than 5 percent of brain microglia each. Their proportion varied by individual and by disease state. One subtype, dubbed cluster 7, expressed numerous genes associated with AD; curiously, it was nearly gone from AD brain.
- Single-cell RNA-Seq reveals nine microglial subtypes in human brain.
- Most microglia are homeostatic, even in AD brain.
- A subtype involved in antigen presentation dwindles in AD.
“The microglia involved in disease are probably in these minor subsets,” De Jager told Alzforum. This implies therapeutic approaches will need to be precisely targeted to affect only the relevant subtype, he added.
Overall, the findings are broadly similar to previous microglial data from both mice and people. They offer a more refined view of different transcriptional states, and they reinforce differences in how humans and mice respond to amyloidosis. Some of the data were presented at a 2018 Keystone Symposium (Jul 2018 conference news).
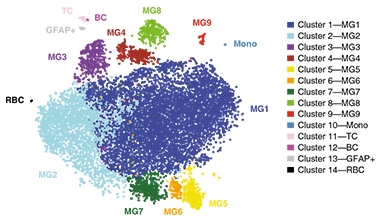
Nine Kinds. scRNA-Seq of human microglia finds that most belong to one of two homeostatic subtypes (MG1 and MG2), and seven other activation states are present in all brain samples examined. [Courtesy of Olah et al., Nature Communications.]
“This is an important resource paper. Every human microglial project is a gold mine,” Oleg Butovsky at Brigham and Women’s Hospital, Boston, told Alzforum. Mark Fiers at KU Leuven, Belgium, agreed. “This dataset will be very valuable to future understanding of the role of microglia in Alzheimer's disease,” he wrote (full comment below).
Most previous studies of gene expression in human microglia relied on bulk tissue samples, and were unable to pick out subtypes (Jun 2017 news; Jul 2017 news; Feb 2018 news). Recently, groups led by Li-Huei Tsai at the Massachusetts Institute of Technology and Marco Colonna at Washington University in St. Louis turned to single-nuclei RNA-Seq of postmortem brain samples to zero in on activation states; alas, the number of microglia isolated from these mixed cortical cell samples was low, around 2,000 to 3,000 (May 2019 news; Jan 2020 news).
In addition, analyzing nuclei rather than whole cells may miss some cytoplasmic transcripts responsible for microglial activation (Oct 2020 news). Whole microglia can only be isolated from living tissue. Just one previous study has done this, when researchers led by Marco Prinz at the University of Freiburg in Germany analyzed 4,400 freshly isolated human microglia by single-cell RNA-Seq (Nov 2019 news).

Expression Profiles. High (red) or low (blue) expression of specific genes characterizes nine microglial subtypes. Circle size indicates the relative percentage of microglia in the cluster that express a given gene, with the largest circle being 100 percent. Ns reflect number of microglia in each cluster. [Courtesy of Olah et al., Nature Communications.]
De Jager and colleagues wanted to examine many more than that. To get living cells, joint first authors Marta Olah and Vilas Menon used fresh autopsy tissue from the dorsolateral prefrontal cortices of 14 participants in the Rush Memory and Aging Project who had had cognitive impairment or AD dementia. Researchers removed tissue samples from the skull within a few hours of death, placing them in a medium that keeps microglia alive (Olah et al., 2018). RNA was extracted within 24 to 48 hours. The authors rounded out this material with surgical resections of temporal cortex from three younger, cognitively healthy people undergoing treatment for epilepsy. From these samples, the authors isolated a total of 16,096 microglia.
Single-cell RNA-Seq defined nine transcriptional subtypes. More than 80 percent of the microglia fell into clusters 1 and 2, which seemed to represent homeostatic cells. These clusters expressed no unusual transcription factors or cell-surface markers. Other subtypes were more distinctive (see image at right). In cluster 3, genes involved in the cellular stress response were active, and these microglia were more abundant in AD brains than in the surgical samples. Cluster 4 turned on genes related to the interferon response, and could be identified by the gene ISG15, which encodes a ubiquitin-like protein induced by interferon. Clusters 5 and 6 sported anti-inflammatory genes, and the transmembrane protein CD83, an immunoglobulin. Cluster 7 expressed antigen-presenting genes and stood out through its high levels of CD74, part of the major histocompatibility complex class II (MHCII). Cluster 8 had no identifying marker, but expressed many metabolic genes. Cluster 9 seemed to contain proliferating microglia, which carried proliferating cell nuclear antigen (PCNA).
The findings dovetail with other microglial studies. Prinz’s whole-cell RNA-Seq analysis of human microglia identified eight of the same clusters, but not cluster 9 proliferating microglia. Data from postmortem brain matches less closely. Both Tsai and Colonna found fewer subtypes, but did implicate interferon in the microglial response to amyloidosis. Colonna reported that microglia in AD brain dial up interferon regulatory factor 8, similar to De Jager’s cluster 4 (Zhou et al., 2020). Meanwhile, in a mouse model, Tsai found an interferon-rich microglial subtype active in the late stage of neurodegeneration. In the same mouse study, Tsai identified a microglial subtype distinguished by MHCII, and a proliferating subtype that appeared early in neurodegeneration (Oct 2017 news).
“It was reassuring that the authors found these same microglial subsets in human brain,” Tsai told Alzforum. She noted that she was unable to identify these subsets in her own snRNA-Seq study of 2,000 microglia from postmortem human brain, probably because of the small number of cells analyzed. Intriguingly, a recent mouse study implicated interferon in the microglial response to tangles, as well (Dec 2020 news).

DAM Bars. The proportion of microglia (green) expressing homologs of mouse DAM genes varies among human clusters. Cluster 7 has the most. [Courtesy of Olah et al., Nature Communications.]
On the other hand, De Jager and colleagues did not find a single microglial subtype that matched the disease-associated microglia (DAM) seen in mouse models of amyloidosis (Jun 2017 news). Rather, DAM genes were highly expressed among four human microglial subtypes: 4, 5, 7, and 8, with cluster 7 the most enriched (see image at left). De Jager noted that amyloidosis in human brain is much slower, and less intense, than in mouse models. “The kinetics are different, and the fundamental microglial biology is probably quite different,” he suggested.
Because cluster 7 expressed many genes associated with Alzheimer’s in addition to the DAM genes, the authors took a closer look at how it changed in AD brain. Immunohistochemistry on sections from eight AD and 11 control brains showed that this microglial subtype made up about 3 percent of all microglia in healthy brain, but only 1 percent in AD. Likewise, an RNA-Seq analysis of bulk postmortem frontal cortex sections from 541 older adults found that only cluster 7 gene expression was dampened in AD brain. Could a dearth of these MHCII-expressing cells hasten AD progression? A recent mouse study hints it could. In it, 5XFAD mice lacking the complex had more plaques and worse inflammation than 5xFAD controls (Mittal et al., 2019).
De Jager is expanding this microglial RNA-Seq data set to delve deeper into specific subtypes. His group has now analyzed more than 200,000 microglia isolated from different brain regions of 75 donors. Samples came from people with different neurological diseases, including AD, multiple sclerosis, frontotemporal dementia, and amyotrophic lateral sclerosis. De Jager believes these data will clarify how microglial activation relates to disease, which could yield clues on how to intervene therapeutically. Because the majority of microglia are homeostatic even in the face of disease, De Jager cautioned against approaches that kill off or suppress microglia en masse (Mar 2018 news). Instead, disease progression might slow if scientists found ways to subtly shift the proportion of microglia in a given activation state.—Madolyn Bowman Rogers
References
News Citations
- A Delicate Frontier: Human Microglia Focus of Attention at Keystone
- What Makes a Microglia? Tales from the Transcriptome
- Human and Mouse Microglia Look Alike, but Age Differently
- Microglial Transcriptome Hints at Shortcomings of AD Model
- When It Comes to Alzheimer’s Disease, Do Human Microglia Even Give a DAM?
- Human and Mouse Microglia React Differently to Amyloid
- Single-nucleus RNA Sequencing Misses Activation of Human Microglia
- The Human Brain Hosts a Menagerie of Microglia
- Changing With the Times: Disease Stage Alters TREM2 Effect on Tau
- Microglia in Tauopathy: Not Just Homeostatic Versus DAM
- Hot DAM: Specific Microglia Engulf Plaques
- Wiping Out Microglia Prevents Neuritic Plaques
Research Models Citations
Paper Citations
- Olah M, Patrick E, Villani AC, Xu J, White CC, Ryan KJ, Piehowski P, Kapasi A, Nejad P, Cimpean M, Connor S, Yung CJ, Frangieh M, McHenry A, Elyaman W, Petyuk V, Schneider JA, Bennett DA, De Jager PL, Bradshaw EM. A transcriptomic atlas of aged human microglia. Nat Commun. 2018 Feb 7;9(1):539. PubMed.
- Zhou Y, Song WM, Andhey PS, Swain A, Levy T, Miller KR, Poliani PL, Cominelli M, Grover S, Gilfillan S, Cella M, Ulland TK, Zaitsev K, Miyashita A, Ikeuchi T, Sainouchi M, Kakita A, Bennett DA, Schneider JA, Nichols MR, Beausoleil SA, Ulrich JD, Holtzman DM, Artyomov MN, Colonna M. Human and mouse single-nucleus transcriptomics reveal TREM2-dependent and TREM2-independent cellular responses in Alzheimer's disease. Nat Med. 2020 Jan;26(1):131-142. Epub 2020 Jan 13 PubMed. Correction.
- Mittal K, Eremenko E, Berner O, Elyahu Y, Strominger I, Apelblat D, Nemirovsky A, Spiegel I, Monsonego A. CD4 T Cells Induce A Subset of MHCII-Expressing Microglia that Attenuates Alzheimer Pathology. iScience. 2019 Jun 28;16:298-311. Epub 2019 May 30 PubMed.
Further Reading
Primary Papers
- Olah M, Menon V, Habib N, Taga MF, Ma Y, Yung CJ, Cimpean M, Khairallah A, Coronas-Samano G, Sankowski R, Grün D, Kroshilina AA, Dionne D, Sarkis RA, Cosgrove GR, Helgager J, Golden JA, Pennell PB, Prinz M, Vonsattel JP, Teich AF, Schneider JA, Bennett DA, Regev A, Elyaman W, Bradshaw EM, De Jager PL. Single cell RNA sequencing of human microglia uncovers a subset associated with Alzheimer's disease. Nat Commun. 2020 Nov 30;11(1):6129. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
KU Leuven
This paper describes the single-cell analysis of microglia from 17 individuals, 10 with AD, four MCI, and three surgical patients, consisting of 16,000 cells, yielding nine distinct microglial clusters. This work is of obvious importance, given the role of microglia in Alzheimer’s and other neurodegenerative diseases.
A strong feature of this paper is that the authors used both postmortem and surgical resections, and subsequently confirmed their findings in a number of third-party datasets by integrating these datasets with their cells. I would be interested to see an even wider integration of human microglial datasets. For example, the authors mention the studies from Del-Aguila et al. and Habib et al., but do not investigate whether these microglia separate into their cluster structure (Del-Aguila et al., 2019; Habib et al., 2017). Also, the inclusion of human microglia from a chimeric mouse model recently published would have been informative (Mancuso et al., 2019). …More
The authors, when comparing results from the surgical resections and postmortem samples, chose to exclude a cluster of cells they associated with cell stress. In agreement with this, we and others have published studies showing that the microglial expression profiles can be influenced by experimental parameters and protocols (Thrupp et al., 2020; Marsh et al., 2020). This might (in part) be a reason for the somewhat limited consistency that most human microglia studies (including this one) find between their own data and other studies.
In conclusion, this dataset will be very valuable for future understanding of the role of microglia in Alzheimer's disease. However, it does remain an open question as to why a number of excellent studies fail to converge on a consistent human-disease-associated microglial profile. Is this due to technicalities? Due to high variation between human samples? Or do microglia play a role much earlier in the pathogenesis of human AD that has been missed?
References:
Sankowski R, Böttcher C, Masuda T, Geirsdottir L, Sagar, Sindram E, Seredenina T, Muhs A, Scheiwe C, Shah MJ, Heiland DH, Schnell O, Grün D, Priller J, Prinz M. Mapping microglia states in the human brain through the integration of high-dimensional techniques. Nat Neurosci. 2019 Nov 18; PubMed. Correction.
Del-Aguila JL, Li Z, Dube U, Mihindukulasuriya KA, Budde JP, Fernandez MV, Ibanez L, Bradley J, Wang F, Bergmann K, Davenport R, Morris JC, Holtzman DM, Perrin RJ, Benitez BA, Dougherty J, Cruchaga C, Harari O. A single-nuclei RNA sequencing study of Mendelian and sporadic AD in the human brain. Alzheimers Res Ther. 2019 Aug 9;11(1):71. PubMed.
Thrupp N, Sala Frigerio C, Wolfs L, Skene NG, Fattorelli N, Poovathingal S, Fourne Y, Matthews PM, Theys T, Mancuso R, de Strooper B, Fiers M. Single-Nucleus RNA-Seq Is Not Suitable for Detection of Microglial Activation Genes in Humans. Cell Rep. 2020 Sep 29;32(13):108189. PubMed.
Habib N, Avraham-Davidi I, Basu A, Burks T, Shekhar K, Hofree M, Choudhury SR, Aguet F, Gelfand E, Ardlie K, Weitz DA, Rozenblatt-Rosen O, Zhang F, Regev A. Massively parallel single-nucleus RNA-seq with DroNc-seq. Nat Methods. 2017 Oct;14(10):955-958. Epub 2017 Aug 28 PubMed.
Mancuso R, Van Den Daele J, Fattorelli N, Wolfs L, Balusu S, Burton O, Liston A, Sierksma A, Fourne Y, Poovathingal S, Arranz-Mendiguren A, Sala Frigerio C, Claes C, Serneels L, Theys T, Perry VH, Verfaillie C, Fiers M, De Strooper B. Stem-cell-derived human microglia transplanted in mouse brain to study human disease. Nat Neurosci. 2019 Dec;22(12):2111-2116. Epub 2019 Oct 28 PubMed.
Marsh SE, Kamath T, Walker AJ, Dissing-Olesen L, Hammond TR, Young AM, Abdulraouf A. Single Cell Sequencing Reveals Glial Specific Responses to Tissue Processing & Enzymatic Dissociation in Mice and Humans. BioRxiv, December 3, 2020
Thrupp N, Sala Frigerio C, Wolfs L, Skene NG, Fattorelli N, Poovathingal S, Fourne Y, Matthews PM, Theys T, Mancuso R, de Strooper B, Fiers M. Single-Nucleus RNA-Seq Is Not Suitable for Detection of Microglial Activation Genes in Humans. Cell Rep. 2020 Sep 29;32(13):108189. PubMed.
Washington University School of Medicine
Washington University in St. Louis
The paper by Olah et al. evaluates the microglial signature through scRNA-seq analyses of live microglia cells from 10 Alzheimer’s disease (AD), four mild cognitive impairment (MCI) and three temporal lobe epilepsy (TLE) patients and compares these data with microglial signatures from two mouse models of neurodegeneration (CKp25 and 5XFAD), as well as with human microglial signatures previously delineated through scRNA-seq and snRNA-seq (Sankowski et al., 2019; Mathys et al., 2019).
The study characterizes four major human microglia populations involved in, respectively, homeostasis, interferon response, antigen presentation, and proliferation. This is in line with the four subsets that were previously found in mouse models of AD. However, the frequencies of the non-homeostatic subsets were significantly reduced in AD/MCI patients compared to those with TLE. Notably, the frequency of the antigen presentation cluster is diminished in AD samples, although immunohistochemical studies have shown an increase in HLA-DR expression. Overall, the functional impact of these microglial subsets remains obscure.…More
Furthermore, Olah et al. showed that the disease-associated microglia (DAM) signature identified in mouse models of AD is not confined to one subset of human microglia, but rather is distributed among all microglia populations, confirming the idea that the mouse DAM signature does not entirely recapitulate the microglial response in human AD. Earlier this year we published a study in Nature Medicine in which we performed snRNA-seq of AD and control patients using brain specimens obtained through ROSMAP. We also reached the conclusion that microglia from human AD patients exhibit a signature partially distinct from that in mouse models: AD-associated risk factors, as well as “homeostatic” genes were upregulated in microglia from AD patients but not in those from controls (Zhou et al., 2020).
Unfortunately, our paper and conclusions have been ignored in the analyses carried out by Olah et al., although the data have been broadly available to the community since February 2020 from the ROSMAP portal. Moreover, the conceptual difference between human AD and mouse models of AD was clearly highlighted in our paper. Our data would have corroborated some of the conclusions reached by Olah et al., but also yielded some important differences that may be due to heterogeneity of patients analyzed, brain regions processed, or technical procedures used to perform snRNA-seq. Evaluation and discussion of these differences would have been helpful for advancing the field of Alzheimer’s disease and neurodegeneration in general.
References:
Sankowski R, Böttcher C, Masuda T, Geirsdottir L, Sagar, Sindram E, Seredenina T, Muhs A, Scheiwe C, Shah MJ, Heiland DH, Schnell O, Grün D, Priller J, Prinz M. Mapping microglia states in the human brain through the integration of high-dimensional techniques. Nat Neurosci. 2019 Nov 18; PubMed. Correction.
Mathys H, Davila-Velderrain J, Peng Z, Gao F, Mohammadi S, Young JZ, Menon M, He L, Abdurrob F, Jiang X, Martorell AJ, Ransohoff RM, Hafler BP, Bennett DA, Kellis M, Tsai LH. Author Correction: Single-cell transcriptomic analysis of Alzheimer's disease. Nature. 2019 Jul;571(7763):E1. PubMed.
Zhou Y, Song WM, Andhey PS, Swain A, Levy T, Miller KR, Poliani PL, Cominelli M, Grover S, Gilfillan S, Cella M, Ulland TK, Zaitsev K, Miyashita A, Ikeuchi T, Sainouchi M, Kakita A, Bennett DA, Schneider JA, Nichols MR, Beausoleil SA, Ulrich JD, Holtzman DM, Artyomov MN, Colonna M. Author Correction: Human and mouse single-nucleus transcriptomics reveal TREM2-dependent and TREM2-independent cellular responses in Alzheimer's disease. Nat Med. 2020 Jun;26(6):981. PubMed.
University of Arkansas for Medical Sciences
Cluster 13 is rather curious. Is there any possibility that scRNA-seq could pick up RNA from cells that microglia have phagocytosed? RNA has the reputation of being exquisitely labile, and I'm sure that's true if your objective is a complete, intact strand. But some of my colleagues are picking up sequence-able RNA from the insoluble aggregates that accrue in the aging brain.
Columbia University Medical Center
Columbia University Irving Medical Scool
We thank our colleagues for their interest in our work and their comments. In a rapidly moving field—such as the study of human microglia in Alzheimer’s disease—it is difficult to have a manuscript fully up to date, in terms of incorporating recently published studies, by the time it is posted online. Drs. Colonna, Fiers, and Zhou highlight excellent recent work that needs to be considered as we interpret our results.
The last major revision of our manuscript occurred at the beginning of 2020, prior to the publication of many of these papers, and they were therefore not incorporated into our article. Our manuscript had a protracted review process, since it was posted publicly on bioRxiv on June 11, 2018 (Olah et. al., 2018). Though the paper has gone through multiple revisions in two different journals since that date, and was presented at several international conferences, the basic message of the study did not change. It includes the discoveries that in the aged brain, the vast majority of microglia are in a state that can be best described as homeostatic, and that one subset (in the bioRxiv version the antigen presentation related subset was named cluster 4) has a negative association with AD dementia. Additionally, we also reported in the preprint that the murine disease-associated microglial (DAM) signature was distributed across a number of human microglia clusters, with the cluster depleted in AD showing the strongest enrichment in DAM genes.…More
Since the observation regarding the prevalence of the homeostatic microglia phenotype in AD, and the discordance between mouse and human in terms of DAM, both represent a major paradigm shift in the field—and accordingly attracted a significant amount of criticism from reviewers—we were delighted to see consistent results replicating our findings emerging in the literature before the printed version of our manuscript became available. In particular, Dr. Colonna and Dr. Zhou’s manuscript is important in this regard, and the observed differences probably arise from the reported reduced ability of single-nucleus RNA sequencing approaches to capture microglial transcriptional activation states when compared to live-cell based approaches (Thrupp et. al., 2020). Larger datasets that are emerging should eventually converge on a more definitive model of microglial population structure that we can apply across studies and platforms, but it is clear that, in humans, there is no single microglial subtype that causes disease.
There are a large number of different transcriptional programs that can be pathogenic in different conditions. In response to Dr. Barger’s comment, we agree that cluster 13 is an intriguing subset of cells that deserve further evaluation. They are quite distinct from the other CD45+ cells that we purified; it is possible that they represent phagocytic cells that have absorbed RNA from other cell types. However, this is speculative at this time, and it is possible that they represent a form of technical artifact. Further work will hopefully provide more answers to this question.
References:
Olah M, Menon V, Habib N, Taga M, Yung C, Cimpean M, Khairalla A, Dionne D, Hopp S, Frosch MP, Hyman BT, Beach TG, Sarkis R, Cosgrove GR, Helgager J, Golden JA, Pennell PB, Schneider JA, Bennett DA, Regev A, Elyaman W, Bradshaw EM, De Jager PL. A single cell-based atlas of human microglial states reveals associations with neurological disorders and histopathological features of the aging brain. bioRχiv. June 11, 2018 BioRxiv.
Thrupp N, Sala Frigerio C, Wolfs L, Skene NG, Fattorelli N, Poovathingal S, Fourne Y, Matthews PM, Theys T, Mancuso R, de Strooper B, Fiers M. Single-Nucleus RNA-Seq Is Not Suitable for Detection of Microglial Activation Genes in Humans. Cell Rep. 2020 Sep 29;32(13):108189. PubMed.
Make a Comment
To make a comment you must login or register.