In April 2006, German-International Alzheimer research strutted its stuff in Bavaria, the state where Alois Alzheimer was born. Scientists met at a secluded lake nestled deep in the Bavarian Alps in the southern part of the state, an area known for its bucolic landscape, skiing, and expressionist art.
Tall Science at Small Retreat: Dispatch from Germany’s Eibsee
Last month at the feet of Germany’s tallest mountain, the Zugspitze, German Alzheimer disease researchers met with colleagues from both sides of the Atlantic to exchange their latest data on research topics ranging from the APP secretases to tau and other topics relevant to AD pathogenesis and therapy development. Having started 6 years ago as the annual retreat for the members of a priority program project by the Deutsche Forschungsgemeinschaft (DFG), the conference drew 90 participants this year. Initiated by Christian Haass of Ludwig-Maximilians-University, Munich, and managed jointly by Haass and Gerd Multhaup at Free University of Berlin, the priority program focused on the cell biology of AD, and has extended government funds to a network of up to 13 German AD labs. Since its inception in 2000, the program has helped broaden the base of AD labs across the country from three major sites that had existed in the late 1990s in Heidelberg, Hamburg, and Munich. By all accounts, the program has increased the visibility of German AD research, in part by supporting young researchers as they set up their own labs, and in part by fostering a German peer group that tries to emphasize a sense of collegiality and cooperation over work in isolation. The program’s reviewers are international. One of them, Peter St. George-Hyslop of the University of Toronto, Canada, went as far as to say that in his estimation, this priority program (or RfA in the U.S.) is the most productive worldwide program working on AD at this time. DFG priority programs expire after 6 years and are not renewable. This one will run out next month, and its history and detailed research results will appear later this year in a special issue of the journal Neurodegenerative Diseases.
Beyond featuring new findings on the γ-secretase complex, the conference showcased a broader cell biology of regulated intramembrane proteolysis by aspartyl proteases. α cleavage of APP emerged from the shadows of the more heavily studied β and γ cleavages, and new studies highlighted its exquisite regulation. APP transport and interactions between amyloid and tau drew attention, as did new mouse models of tau and mutant APP knock-in mice. Scientists advanced new modulators of amyloid β’s effect on synaptic function, new ways of studying Ab oligomers, and discussed the interplay between lipids and APP processing. Others told of the search for better NSAIDs, and offered the latest update on the Swiss patients in Elan’s aborted AN1792 immunotherapy trial.
Before the science started, however, Haass introduced a new prize in the field. The German Hans and Ilse Breuer Stiftung is a family foundation that has agreed to take over funding the Eibsee conference as an annual platform for German AD research. The foundation also committed funds for graduate student stipends and for an annual “Alzheimer-Forschungspreis.” Selected by an international jury, the prize rewards a German AD researcher with $100,000 of unrestricted research money. Its size equals that of the Potamkin Prize awarded since 1988 at the annual meeting of the American Academy of Neurology. The Potamkin Prize arguably has become the most prestigious award for AD research in the U.S. and is funded by a family foundation in memory of Luba Potamkin, a businesswoman diagnosed with dementia in 1978. Similarly, the Breuers decided to do something about Alzheimer disease after watching the suffering of their mother Ilse. The inaugural prize went to Harald Steiner, at the LMU, for his research on γ-secretase. (Steiner is a trainee of Haass’, who noted that he abstained from the prize selection process.) In Germany, people still look mostly to the government for research funding, and the scientists at the conference clearly hoped that the Breuers’ lead would inspire additional private philanthropy in their country.
Below are excerpts of the presentations and discussions:
Synapses: Duress and Relief
After recapping some milestones in AD research, Dennis Selkoe of Brigham and Women’s Hospital, Boston, started off the science by saying that a key challenge in current AD research has become to understand precisely what it is that subverts the function of certain synapses at the earliest stage of disease. Unsurprisingly for followers of the field, Selkoe suspects that Aβ oligomers, flowing in interstitial spaces, reach the synaptic cleft and interact with components of the postsynaptic machinery to throw off synaptic transmission and start disease cascades. The Alzforum has followed the evolving story of how preparations enriched for Aβ oligomers are thought to disrupt LTP in cultured brain slices and interfere with learning (Wang et al., 2002; Walsh et al., 2002; Cleary et al., 2005; Townsend et al., 2006). New developments reported at the Eibsee concern ongoing confocal microscopy studies that aim to visualize what oligomers do to dendritic spines of fluorescently labeled neurons. For example, Ganesh Shankar, working with Selkoe and Bernardo Sabatini at Harvard Medical School, is finding that picomolar quantities of secreted oligomer fractions separated by size exclusion chromatography cause a reduction in the density of spines on dendrites of cultured rat hippocampal slices. The dendrites stayed in place throughout the experiment but were no longer studded with spines a few days after oligomer application. Spine numbers began to drop the day after oligomer exposure and recovered once the oligomers were washed out, Selkoe said. Anti-Aβ antibodies restored spine number, he added, much like they appear to rescue previously observed oligomer effects on LTP in rats (Klyubin et al., 2005).
This is among the first studies to note how spines wither under direct exposure of Aβ by using a combination of diOlistic “gene gun” labeling and confocal microscopy. Other groups are using these methods to examine spines in transgenic mice. Researchers led by Floyd Bloom are noticing spine loss early on in the dentate gyrus of PDAPP mice (see SfN conference story) and similar results using Golgi impregnation are published (see Jacobsen et al., 2006). While imaging hippocampal slices of two different mutant APP- and PS1 transgenic mouse strains, scientists led by Michael Shelanski at Columbia University, New York, also noticed a dearth of spines, as well as swollen dendrites (Moolman et al., 2004). The spines in the amyloid transgenic strains looked different, with longer necks and larger heads, Shelanski said. Dendritic spines look similarly altered in some mouse models of inherited mental retardation. At the Eibsee, Shelanski showed diOlistic staining of spines of an AD patient and an 89-year-old person who had died after aging normally. (This kind of spine imaging works only in freshly fixed tissue, not in stored samples from brain banks that have been in fixatives for long periods of time, Shelanski noted.) The AD patient appeared only to have had about half of the dendritic area and number of spines as did the control, suggesting that, in this regard, the transgenic mice model AD reasonably well.
Shelanski reported on his group’s search for agents that might reverse the detrimental effects of Aβ oligomers on hippocampal synapses. He believes that factors that boost signaling through the cyclic AMP-PKA CREB signaling pathway could do the trick. (Pioneered by Eric Kandel, this pathway is thought to underlie memory processes.) Prior work had shown that rolipram—an inhibitor of phosphodiesterase 4 (PDE4) that prevents breakdown of cyclic AMP—can improve some forms of memory in mice (Barad et al., 1998). Rolipram also reverses the LTP deficit in hippocampal slices, as well as learning and memory deficits of APP/PS1 transgenic mice (Gong et al., 2004). Interestingly, once the mice had received a course of the drug, they kept performing better for several months without further treatment, as if the CREB pathway had been reset for the long term, Shelanski noted. However, efforts in biotechnology companies to bring new PDE4 inhibitors without the gastrointestinal side effects of rolipram to market have not yet succeeded.
Shelanski’s newer work takes a different tack toward the same goal of counteracting Aβ’s effect on synaptic function. It focuses on proteasome degradation, and specifically on ubiquitin C-terminal hydrolase-L1 (UCH-L1). This neuronal enzyme recycles ubiquitin, making it available for re-use in further cycles of tagging and targeted degradation of waste proteins in the protein grinder. UCH-L1 is known for its association with rare cases of Parkinson disease (e.g., Lincoln et al., 1999). It rarely comes up in AD research, except in one study that reported a down-regulation in brains of sporadic AD cases (Choi et al., 2004). Other data have suggested that it improves LTP induction in the sea slug Aplysia.
This and other hints led Shelanski’s team to develop a membrane-permeable version of UCH-L1 that enabled them to manipulate its concentration inside neurons. Called TAT-HA-UCH-L1, the construct exploits an HIV protein that induces micropinocytosis at lipid rafts and thus ferries the enzyme across the cell membrane. Much like rolipram, TAT-HA-UCH-L1 restored LTP in young APP/PS1 double-transgenic mice, Shelanski reported. It also normalized basal synaptic transmission in older transgenic mice and rescued the spine loss in their hippocampal slices, as well as in Aβ-treated slices of wild-type rodents. The reverse was also true; that is, Aβ interfered with the activity of UCH-L1 and reduced availability of mono-ubiquitin.
How would Aβ interact with UCH-L1? Shelanski believes it does so indirectly, through signaling from the outside. In any event, the added UCH-L1 propped up proteasome function either by freeing up more mono-ubiquitin, or perhaps by dimerizing and acting as a ubiquitin ligase (Liu et al., 2002). On this question, Shelanski suspects primarily the first function, since a mutation in the UCH-L1 hydrolase domain abolished its ability to restore LTP. Boosting UCH-L1 function could serve as a therapy in its own right or together with amyloid-lowering approaches, Shelanski noted. The TAT-HA-UCH-L1 construct itself won’t become a drug because it would require injection and, as a protein, could trigger an unwanted antibody response. Instead, this research serves to inform the search for small molecules, Shelanski said.—Gabrielle Strobel.
Two speakers, Charles Glabe and Mathias Jucker, focused on early aspects of Aβ aggregation. Glabe, of University of California, Irvine, proposed that oligomers of proteins with different amino acid sequences—and linked to a range of amyloid diseases—share not only common structures and pathways of aggregation. They also wreak havoc in basically the same way; that is, they likely assemble into pore-like structures inside membranes (Glabe, 2006; Glabe and Kayed, 2006; see also ARF Live Discussion on amyloid channels). A lesson emerging from Glabe’s newest research is that prefibrillar oligomers are not a required step toward fibril formation. In other words, the classic pathway of monomer-dimer-oligomer-protofibril-fibril-plaque is but one of several modes of aggregation; at least one other pathway exists beside it.
Glabe noted that evidence pointing to oligomeric forms of amyloid proteins as the primary toxic species in their respective disease is growing. Even so, researchers continue to wrestle with the technical challenges of distinguishing one type of oligomer from another, and from monomer and fibrils, for that matter, in tissues containing complex mixtures of the many incarnations of a given protein.
Glabe’s laboratory uses conformation-specific antibodies to pick these forms apart, and new ones the group has generated are proving useful in describing one such alternate aggregation pathway. Their previous antibody, A11, recognizes prefibrillar oligomers, not monomer or fibrils, of a growing number of amyloidogenic proteins, 24 to date (Kayed et al., 2003; Luibl et al., 2006; Sanbe et al., 2005). Its structural antigen appears to be a precursor of a later aggregate, an annular form that makes troublesome pores in membranes much in the way Hilal Lashuel and Peter Lansbury have proposed (Lashuel et al., 2002). These ring-shaped protofibrils are what the new antibody recognizes, and it does so, again, for a range of proteins of different amino acid sequences, Glabe said. The lab dubbed this new polyclonal “Officer,” a playful reference to the American policeman’s stereotypical fondness for donuts. (The patrol cars frequently seen parked outside donut shops don’t reflect elevated crime rates in those establishments, but in the minds of Glabe’s group, the Officer antibody might yet pull a perpetrator of a different sort.)
Separate aggregation pathways imply that a monomer of, for example, Aβ or α-synuclein, can undergo at least two distinct conformation changes. One would lead it down the classic pathway of forming a fibril nucleus and then fibrils. The other conformation would lead via A11-positive prefibrillar nuclei, which in the atomic force microscope look like spherical blobs, to the Officer-positive protofibrils, which appear as smooth rings. Preliminary data suggest that immunoprecipitation with the Officer antibody isolates pore-like structures from human AD brain but not control, Glabe said. The A11 and Officer antibodies do not stain amyloid deposits in AD or, for example, diabetes tissue, but a third antibody that is specific for a generic fibrillar epitope does. Called OC, its tissue staining is particularly stark as it comes up blank in control tissue. “This is purely a pathogenic epitope,” Glabe said. OC reaction with brain extracts correlates with the affected patient’s MMSE rating before death, but A11 does not. In LaFerla’s triple-transgenic mice, the OC antibody recognizes certain intraneuronal Aβ deposits associated with autophagic vesicles.
Exposing Aβ prefibrillar oligomers to liposomes reduces their A11 staining but brings up Officer staining, suggesting that lipids enable the prefibrillar oligomer to assemble into pores. Taken together, ongoing work suggests that A11 recognizes a misfolded intermediate on the way to pore formation. With pores poking tiny holes in their membranes and changing the cell’s permeability to various agents, all cells will have trouble maintaining their ion homeostasis and metal-dependent chemistry such as antioxidant processes, said Glabe, but none more so than neurons, which depend critically on functional neurotransmission (Kayed et al., 2004). Finally, Glabe added, comparing the structural antigens for these three antibodies has taught the scientists that size does not indicate conformation. Some prefibrillar and fibrillar oligomers have the same size but distinct conformations, and they will go down separate aggregation pathways.
Mathias Jucker, of the University of Tuebingen, Germany, expanded on an ongoing study aimed at identifying the particular form of Aβ that is able to “seed” amyloid deposition. For a summary of this line of research, see prior ARF conference report. Jucker asked what it is about Aβ’s conformation, or cofactors imparting that conformation, that enables some forms of Aβ to seed deposition in a tissue that is ripe with rising Aβ accumulation but still short of laying down actual amyloidosis itself. Jucker has searched for the winning concoction by testing a range of different forms of Aβ. Since the prior report, preparations by John Fryer and David Holtzman, Gerd Multhaup, Rakez Kayed and Charlie Glabe, Uli Ebert and Heinz Hillen all came up negative. Likewise, the cell-secreted oligomers developed by Dominic Walsh, and used in LTP, behavior, and spine imaging studies, have not, so far, seeded deposition in APP-transgenic backgrounds, although it should be noted that the scientists have not so far concentrated the oligomers to quite the level where they would expect to see seeding occur. (The newest contender, Karen Ashe's Aβ*56, has not been tested.) By contrast, AD brain extract seeded powerfully in Lary Walker’s hand, as do various AD-transgenic extracts in Jucker’s (Walker et al., 2002).—Gabrielle Strobel.
γ-Secretase: Ins and Outs of a Voracious Membrane Protein Grinder
Several researchers reported their latest data on the γ-secretase complex and APP processing. This research has expanded greatly beyond an initially narrow focus on Aβ production to where it now represents a lively subarea of cell biology that explores how aspartyl proteases carrying a GxGD motif carry out regulated intramembraneous proteolysis (RIP) of type 1, or single-pass, membrane proteins. γ-secretase remains the most famous representative of this group of proteases, but the related, and somewhat simpler, family of signal peptide peptidases (SPPs) is becoming better known and offers useful comparisons for the study of γ-secretase. New trends in this area include a growing realization that γ-secretases and SPPs are using a similar catalytic mechanism, whereby multiple cleavages inside the membrane give rise to a multitude of different fragments, Aβ40 and 42 being but two of them. This mechanism is not processive like that of a peptidase, which chops down its substrate from one end; rather, the substrate moves through the membrane stepwise and gets cleaved in particular positions.
Researchers are beginning to view γ-secretase as a membrane proteasome, or even as a sort of kitchen sink garbage disposal, whose job it is to remove residual transmembrane stubs of many different membrane receptors and proteins, such as APP. In parallel to that activity, though, γ-secretase also carries out highly specific cuts necessary for nuclear signaling. As the list of γ-secretase substrates continues to grow past the 25 or so known to date, researchers are realizing that this protease cares less about the amino acid sequence of its substrates than typical proteases do. Rather, it primarily requires that the previous step of α or β cleavage, also called ectodomain shedding, generate a fragment of a particular length that sports a free N-terminal. Beyond that, the question of how substrate specificity is conferred remains a puzzle.
Another big question concerns regulation. To some investigators, γ-secretase and SPPs look to be rather indiscriminate beasts that cut constitutively once suitable substrates are present. To them, most of the finely tuned regulation and important biology appears to occur upstream, at the level of α cleavage. That said, a brand new secretase complex member and ongoing work on lipids suggest that there is much more to γ-secretase modulation than presently meets the eye.
Finally, one emerging theme concerns comparisons with channel proteins. First glimpses of the complex’s putative 3D structure, and cell biology studies of the way it assembles, are reminding scientists of known aspects of ion channels or the sec61 protein pore through which type 1 membrane proteins slide on their way off the ribosome and into the ER. (Truth be told, this notion has been around for a while; see, e.g., Fred Van Leuven’s comment, and the idea is now gaining new data.) The Eibsee conference offered news on all these fronts. Below find presentation summaries.
Gang Yu, of the University of Texas Southwestern Medical Center in Dallas, started things off by recapping his recent paper showing that γ-secretase action begins when the complex member nicastrin recognizes the substrate (see news and discussion at Shah et al., 2005). Yu found that α or β cleavage of a given substrate protein, for example, APP or Notch, will expose a membrane-bound stub of usually less than 100 amino acids with a new free N-terminus. (In the confusing terminology of RIP, these stubs are called C99/C100/βCTF for APP, NEXT for Notch.) It is this stub that nicastrin latches onto and then swivels around, a bit like a construction crane, to move the substrate deeper into the γ-secretase complex for docking and cutting. Nicastrin forwards the substrate in this way by way of its ectodomain, whereas its transmembrane domain plays a role in the assembly and the activity of the γ-secretase complex. (see image below). Importantly, this crane-like ectodomain of nicastrin appears to select potential substrates more by their length and the presence of the free N-terminal stub than by their sequence, Yu said. Nicastrin uses a conserved DAP domain and, in particular, a glutamate at amino acid position 333 to interact with the substrate, and this site on nicastrin is indeed required for γ-secretase activity. In studying nicastrin’s modus operandi, Yu noted some similarities to how amino peptidases function. These are enzymes that catalyze the hydrolysis of the terminal peptide bond at the amino end of a polypeptide. This study proposes a common mechanism for how nicastrin recognizes all γ-secretase substrates, but it remains to be experimentally tested with regard to most substrates besides APP and Notch. The reality will likely prove to be more complex than this current model, Yu noted.
Nicastrin, via its ectodomain, recognizes the new free N-terminus of the substrates (and via its TM domain, participates in γ-secretase assembly and activity) [Figure courtesy of Gang Yu]
Harald Steiner, of Ludwig Maximilians University in Munich, first recapped some milestones in γ-secretase research, including its reconstitution in yeast (Edbauer et al., 2003) and, since then, in various other cell types. Later, it turned out that distinct γ-secretase complexes exist, made up by different combinations of the presenilin and Aph isoforms plus nicastrin and Pen-2 (Shirotani et al, 2004; Serneels et al., 2005). A pressing goal at this point is to find out exactly how the enzyme complex distinguishes APP from Notch (and other substrates) so that this information can help guide the search for new γ-secretase modulator drugs that can tweak APP cleavage selectively. At the Eibsee conference, talks and informal discussion suggested that several competing groups appear to be converging on the same spot on the protease. In his keynote lecture, Selkoe recounted how his group’s pursuit of this site developed from follow-up studies of Paul Greengard’s discovery that the cancer drug Gleevec inhibits cleavage of C100 (the APP substrate generated by BACE cleavage) but not of the corresponding Notch substrate (Netzer et al., 2003). Certain kinase inhibitors turned out to do the same thing, as does ATP, and these compounds all appear to act on a nucleotide-binding site on γ-secretase (Fraering et al., 2005). Michael Wolfe, at Brigham Hospital, has proposed a site for substrate docking that lies near the active site. It might serve as a stopover between the substrate’s initial binding to nicastrin at the surface of the protease and its eventual cleavage deep inside the protease (Kornilova et al., 2005).
For his part, Steiner is zeroing in on the same site. Coming originally from an interest in how the substrate transits from nicastrin to the active site, Steiner, Haass, and colleagues are finding that substrate docking and distinguishing between APP and Notch may occur in that very same place near the active site. When the scientists took an evolutionarily distant, roundworm cousin of γ-secretase and swapped its active site with that of presenilin-1, the chimera processed APP just fine but could not cut Notch. In-vivo tests in a C. elegans Notch mutant confirmed that something about this presenilin-1 active site was specific to APP. Mutation in its transmembrane domain 7 revealed this something to be a single amino acid, namely a leucine at position 383. This residue represents the X position of a GXGD motif on this domain that lies next to the catalytic site. Steiner’s work suggests that the final substrate selection happens there, as Wolfe had proposed. In effect, the GXGD motif might be a final “gate” to the active site, Steiner said (Yamasaki et al., 2006). The motif occurs in γ-secretase and its relatives, the signal peptide peptidases, and its “D” actually represents one of the two proposed catalytic aspartates, implying that GXGD encompasses both substrate selection and part of the active site. Its role, if any, in the selection of other γ-secretase substrates is not clear yet.
In the discussion, Selkoe speculated that a Walker-like motif exists on transmembrane domain 7 of presenilin that might account for ATP binding and represent a site of allosteric regulation of γ-secretase. Described by John E. Walker as sequence motifs indicating the presence of nucleotide binding domains, these sequences typically occur in protein regions accessible to water so they can support ATP hydrolysis. It is unusual for such a motif to lie hidden inside a hydrophobic region of an intramembranous enzyme complex, but then, most everything about γ-secretase is unusual. The presence of such a motif might yield clues to γ-secretase function, that is, where water enters into the lipid bilayer to enable breaking of the substrate’s peptide bonds, and how the catalytic aspartates in presenilin’s transmembrane domains become protonated in preparation for proteolysis, Selkoe noted.
This Walker-like motif showed up in yet another guise. By studying APP dimerization, Gerd Multhaup arrived at this region, as well. With postdoc Lisa Munter, Multhaup discovered that it turns out to harbor a third, potentially critical dimerization spot on APP that influences the sequential cleavages of the γ-secretase substrate. By doing so, it helps determine what forms of Aβ come out of this process.
APP dimerization itself, by two prior points of contact between monomers, had been published before (see, for example, Beher et al., 1996; Scheuermann et al., 2001). Some other γ-secretase substrates are thought to dimerize, as well. At the conference, Multhaup started out by saying that his group had discovered a third, new site of contact between two APP molecules (as well as other substrates) that mediates dimerization. A FRET study confirmed dimerization in living cells. On APP, this new site includes three consecutive GxxxG motifs. From other research, such motifs are known to promote interaction of protein helices, and a recent Alzforum Discussion challenged the field to ponder whether they somehow might mediate Aβ toxicity. On APP, one of the GxxxG motifs within the Aβ sequence represents a particularly important site for APP dimerization and lies within the Walker-like motif. Multhaup speculated further that the generation of Aβ42 depends on the GxxxG motifs by influencing APP-APP interaction. Further details on this work will be reported upon publication.—Gabrielle Strobel.
Mechanistic studies of γ-secretase, and the search for good modulators, would make a leap forward if the scientists could simply look at the protease and see its structure. With at least 18 transmembrane domains, the complex is too fiendishly complicated to have yielded to x-ray crystallography to date, and in the meantime, some groups are making do with electron microscopy. At the Eibsee, Dennis Selkoe described one of the first such attempts, performed with collaborator Huilin Li at Brookhaven National Laboratory in Upton, New Jersey, to use the synchrotron there to increase conventional EM resolution. The work just appeared in PNAS online (Lazarov et al., 2006; see comment by Boris Schmidt).
Joint first authors Vlado Lazarov of Brookhaven and Patrick Fraering of Selkoe’s group describe how they purified a cell-derived human γ-secretase complex, assured its proper composition and activity, labeled the nicastrin ectodomain with a lectin to orient the particles, and then developed averaged EM images as well as 3D surface reconstructions. The images show a slightly elongated particle about 12 nm across, with irregular bulges and indentations. At first glance, it resembles an ion channel because it features a hollow central chamber. This could shield the active site needed for hydrolysis from the hydrophobic environment of the membrane. The particle also has 2 nm-wide openings, one at the top and one at the bottom. Selkoe showed a movie of the particle (download at PNAS website), as well as a silhouette model of its surface contours into which the scientists had slotted its component proteins in a first-pass attempt of depicting how they might fit together. (Efforts to improve on these first images with cryoEM are ongoing, Selkoe noted.)
At the conference, a lively discussion ensued. One question concerned how the substrate would access the active site in the central chamber. It cannot flow in from the top or bottom, as is the case with ion channels, but must enter laterally from the lipid bilayer through an opening in the side wall, or some sort of “gating” mechanism. This initial EM study shows the γ-secretase complex in a closed form and cannot answer that question. Another question is where water would enter and the reaction products leave. Selkoe suggested that the two small holes might serve as conduits toward the extra- and intracellular compartments, where, in the case of APP, Aβ could exit through one hole and AICD through another. A third question was what additional proteins, or allosteric interactions, enabled the complex to plug up its central chamber and prevent water flooding in, which would kill the cell. Tom Rapoport’s seminal paper showing the structure of the sec61 protein channel featured a plug that seals the pore in the default state so the ER won’t collapse. It also proposes a mechanism for opening a lateral gate, (Van den Berg et al., 2004). For γ-secretase, the nicastrin ectodomain is one obvious candidate, Selkoe noted. Additional, regulatory, complex members are just beginning to be discovered (see St George-Hyslop talk below).
Haass’ group is approaching these questions with ongoing structural work using presenilin homologs from the SPP family, which have cropped up in database searches. Functioning as dimers and trimers, they are easier to study. Regina Fluhrer (who did double duty as conference organizer and presenter) presented data showing that, enzymatically, the active site of these SPP-like proteins (SPPLs) works just like that of γ-secretase. It uses the same cleavage mechanism even though its members do not form large complexes with nicastrin, Aph-1, and Pen-2. This suggests that such a complex may not be necessary for the formation—and visualization—of the central pore. A recent paper characterized the intracellular location and mutant phenotype of some of those SPPLs (Krawitz et al., 2005). At the Eibsee, Fluhrer showed new data suggesting that SPPLs tend to process type 2 transmembrane proteins, such as TNFα, whereas presenilins cleave type 1 transmembrane proteins (Fluhrer et al., 2006, in press). Type 2 transmembrane proteins are similar to the type 1 class in that they span the membrane once, but they run in the opposite orientation; that is, the amino-terminal side of their transmembrane domain reaches into the cytoplasmic side of the membrane and their COOH-terminal portion is exposed to the exterior. (Type 3 proteins span the membrane multiple times, like presenilin itself.) In other words, SPPLs act like “inside-out” presenilins, using the same enzymatic mechanism from an inverse orientation inside the membrane (see image by Haass et al. below).
When the Haass, Selkoe, and Takeshi Iwatsubo labs each reconstituted a functioning γ-secretase complex within a few months of each other, they learned that presenilin, nicastrin, Aph-1, and Pen-2 were the bare-bones requirement for activity in vitro. Yet in vivo, the complexes are larger, varied, and bound to be tweaked by a number of additional proteins. At the meeting, Peter St George-Hyslop introduced a new player in this area when he reported that his group had identified a fifth—and regulatory—complex component called TMP21. Itself a type 1 transmembrane protein (though not a γ-secretase substrate), TMP21 was previously identified as a member of a cargo protein family involved in proofreading material that is being shipped through the endoplasmic reticulum (ER) and trans-Golgi network (TGN) to the cell surface. Intriguingly, the TMP21 gene maps to chromosome 14 in a cluster including PS1. This cluster is conserved down to fugu (a poisonous Japanese pufferfish that is becoming a staple of comparative genomics), suggesting that transcription of these genes and their function might be conjoined.
St George-Hyslop’s team showed that besides being part of the p24 cargo complex, TMP21 also occurs in several different presenilin complexes, in which it functions quite differently. It selectively restrains γ cleavage (which generates Aβ from APP C100, for example, and Nβ from NEXT) but has no effect on ε cleavage (which generates the AICD product from C100, and NICD from NEXT). Removing TMP21 from the complex by siRNA increased Aβ/Nβ production but left AICD/NICD production unchanged. TMP21 binding to γ-secretase is highly specific, and its modulation of the γ cleavage is not due to a trafficking or assembly glitch but likely represents a direct interaction, St George-Hyslop said.
By identifying the first of what is likely to be a growing number of γ-secretase modulators, this work offers a fresh glimpse at the properties of the massive, 650 kDa γ-secretase, St. George Hyslop said. For example, the ligand-dependent ε cleavage, which is so crucial for signal transduction to the nucleus in embryonic and adult life, might well be under quite separate control from the γ cleavage, which is increasingly being associated with membrane “garbage” removal (Kopan and Ilagan, 2004). This is new because until recently, researchers have tended to view the different cleavages on nearby spots of a given substrate not as distinct activities but as more likely reflecting a single, loosely defined property. The work also offers a new approach for ongoing efforts to find therapeutic γ-secretase modulators. Rather than pointing to a way to distinguish between APP and Notch substrates per se, small-molecule TMP21 mimics might inhibit Aβ (and Nβ) production while sparing NICD and its signaling functions. This paper appeared last week in Nature (Chen et al., 2006).—Gabrielle Strobel.
Assembly, Traffic Escorts, Fats All Control APP Processing
Scientists believe that of the different γ complexes that exist in vivo, some are tuned for Notch, some for APP, some to yet other substrates. This tuning probably happens during the assembly phase, implying that there must be regulation at this level. Christoph Kaether, formerly of Haass’ group and now at the Leibniz Institute for Age Research in Jena, Germany, studies where in the cell the complex is put together, and what other proteins control this process. Prior work in this area has established that the complex fully assembles in the ER, where Aph-1 and nicastrin first get together, stabilize presenilin, and finally are joined by Pen-2. The complete complex then leaves the ER, moving through the TGN and up to the cell membrane, where it later becomes endocytosed and degraded. A recent study answered, at least in cultured cells, if not yet for neurons in vivo, the longstanding debate about where in the cell γ-secretase is active. It showed that the complex does not begin to be active until it has reached the cell membrane. Cleavage then occurs there, in endosomes, and also in lysosomes, Kaether said (Kaether et al., 2006). This finding ties into an emerging line of thought about dysregulation of endocytosis and autophagy in early AD.
Assembly in the ER is a characteristic the γ-secretase complex has in common with channels, Kaether said. They, too, are made up of subunits that each have an ER retention signal on them, and this signal becomes masked once the subunits have clicked together and are ready to “move up” to the plasma membrane. Surface proteins such as the acetylcholine receptor and B and T cell receptors also fall into this class.
At the Eibsee, Kaether outlined new work detailing retention/retrieval signals in the complex components, as well as additional proteins, which together govern this process. Kaether found two such signals, one in PS1’s C-terminus and one in Pen-2’s first transmembrane domain. A trafficking protein recognizing these signals might be Rer1, the human homolog of a small yeast protein known to retain unassembled proteins in the ER (Sato et al., 2003). Kaether and colleagues found that human Rer1 resides in ER and Golgi, where it binds to the retention signal in Pen-2. Rer1 limits the rate of γ-secretase assembly. The scientists propose that Rer1 cycles between the ER and the Golgi, where it traps unassembled Pen-2 that has escaped from the ER and returns it. Completion of the complex masks the Pen-2 retention signal, and the complex can slip past Rer1’s surveillance in the Golgi. (For more on APP trafficking, see Walter and Lichtenthaler talks.)
Like the γ-secretase components, APP percolates from the ER through the TGN and to the plasma membrane, where it later undergoes endocytosis and degradation in lysosomes. Researchers think that, at least in experimental cell models, APP is subject to α cleavage while in the secretory half of this life cycle and to β and γ cleavage while at the cell membrane and in endosomes, but they do not know how membrane lipids govern this segregation. Cholesterol is most often invoked, but at the Eibsee conference, Jochen Walter of the University of Bonn presented his work on glycosphingolipids. Conventional wisdom has viewed these membrane components as merely structural, but that is changing. Cells make different varieties of these complex molecules by taking ceramide and icing it with various sugars in the ER/Golgi compartments. Their transport through the secretory pathway closely tracks that of APP. Glycosphingolipids further intrigue scientists because neurons contain lots of them, preferentially in detergent-resistant membranes (DRMs, also called lipid rafts), and their levels change in AD and in other neurodegenerative diseases.
In earlier work, Walter inhibited the addition of glucose to ceramide, that is, the first step in glycosphingolipid synthesis. In its absence, cellular expression of APP stayed the same but secretion of both APP and Aβ dropped. Pulse-chase experiments showed that the reason lay in defective maturation, stability, and transport to the cell surface of APP (Tamboli et al., 2005). Since then, RNAi suppression of the enzyme catalyzing the second step in glycosphingolipid biosynthesis, as well as other experiments, has produced similar results in support of the hypothesis that these lipids aid APP maturation and trafficking, Walter said. This may well affect APP processing, as well, Walter noted. Experiments with particular sphingolipids, such as purified gangliosides, suggested that they can promote Aβ generation.
A second line of research in Walter’s lab explores the intraneuronal trafficking of BACE. (Besides Walter’s research, BACE drew surprisingly little attention at a conference otherwise heavily weighted toward APP processing.) During his time in Christian Haass’ lab, Walter had linked phosphorylation to BACE trafficking. Like presenilin, BACE becomes reinternalized into endosomes at the cell membrane. However, rather than being subsequently degraded, non-phosphorylated BACE molecules recycle directly up to the cell surface again, whereas a phosphorylated form of BACE1 journeys backwards through the TGN (Walter et al., 2001; Wahle et al., 2005). This phosphorylation-dependent retrograde transport of BACE1 to the TGN is mediated by sorting proteins that go by a mouthful of a name, that is, Golgi-localized, γ ear-containing, ADP ribosylation factor-binding, or GGA proteins, for short (Bonifacino, 2004). To read up on the emerging area of BACE recycling, see also He et al., 2003; ; and Scott Small’s recent study on retromer proteins.
Can this BACE trafficking change APP processing? There is no answer yet to this question, but Walter believes that GGA1 might hold a clue because these proteins are implicated in two-way transport of cargo proteins through the secretory pathway. Walter’s ongoing work focuses on GGA1, a neuronal variant whose levels he found to be decreased in brain lysate of AD patients. Reducing GGA1 function experimentally caused BACE1 to pile up in the endosomal compartment, Walter noted, where other studies have begun noticing it, as well. APP is a minor one among BACE1’s many substrates, yet scientists believe this cleavage occurs at least in part in the endosomal compartment. Adding to the intrigue around GGA proteins is the finding that they affect trafficking of SorLa, another sorting protein suspected of influencing APP processing (Andersen et al., 2005).
That lipids affect APP processing is quite widely accepted, yet the opposite—that Aβ affects lipids—is a newer, more provocative idea. Tobias Hartmann reviewed a recent discovery and added new twists for further discussion. Last November, Hartmann’s group proposed a set of interlocking feedback loops connecting γ-secretase activity and membrane lipid production, (see news summary and schematic drawing). In brief, Hartmann suggests that not only do cellular lipids such as cholesterol and sphingomyelin regulate γ-secretase activity, but also Aβ itself in turn regulates levels of those lipids. This is specific in that the products of γ-secretase substrates other than APP do not. The gist of the proposed regulatory pathways is that Aβ42 performs the normal, physiologic function of spurring the activity of the degradative enzyme sphingomyelinase, in essence lowering sphingomyelin levels. The ensuing decrease in sphingomyelin would then lift a block on γ-secretase function. For its part, Aβ40 at physiological concentration is reported to keep cholesterol levels low by blocking synthesis at the same HMG-CoA reductase step that statin drugs inhibit. This second cycle closes, as decreasing cholesterol concentrations would end this lipid’s stimulatory effect on γ-secretase activity.
This work has created a buzz in the field, and at the Eibsee, Hartmann added further twists. One concerns the question of how this interdependent system would react to FAD mutations. Hartmann noted that a growing body of data finds a strong correlation between the Aβ42/40 ratio and when disease begins. Specifically, mean age of onset tends to be lower in people who had less CSF Aβ40 and more Aβ42 for a set of FAD mutants Hartmann’s group examined. This change in the ratio would effectively uncouple a formerly integrated regulation of lipid levels: Aβ40 normally blocks HMG-CoA reductase and FAD mutations tend to weaken that inhibition, leaving the cells with more cholesterol. Aβ42 normally activates SMase and many FAD mutations enhance it, leaving the cell with less sphingomyelin and more of its metabolite ceramide, which has been linked to neuronal apoptosis. This data appear to fit with Kumar-Singh et al. (2005), and Wang et al. (2006), who suggest that Aβ40 might have a protective function that falls away in Aβ40-lowering FAD mutations (see recent Alzforum Discussion).
Exactly how might Aβ control lipid levels? Hartmann said that Aβ42 acts directly on sphingomyelinase, but how Aβ40 might inhibit HMG-CoA reductase remains a mystery. The peptide does not act on the gene’s promoter. Cells keep Aβ40 away from the enzyme’s site of residence in the endoplasmic reticulum, so it can’t be a direct action but likely requires a messenger in between. Additional challenges in this line of investigation include human testing, for example, by measuring sphingomyelinase activity and cholesterol levels in people with FAD mutations, and assessing the effect of related peptides, such as Aβ38, on cellular lipids.—Gabrielle Strobel.
Of the three major APP processing steps, the richest and most varied biology appears to lie ahead for investigators who probe α cleavage. Long dismissed as “the other” cleavage that precluded Aβ formation, it languished in the shadows while the field trained its glare squarely on Aβ.
γ cleavage was the α male of the pack and, to be sure, tweaking it to shift the Aβ42 ratio remains a main focus in AD drug development. Even so, academic research on α-secretase is beginning to flourish. Below are summaries of five Eibsee talks on different angles of its biology. Paul Saftig, at Christian-Albrechts University of Kiel, Germany, introduced the topic broadly by noting that α cleavage, also called ectodomain shedding, is proving to be a critical step in the function of a wide range of a cell’s surface molecules. Scientists to date have identified about 40 different sheddases, or ADAMs. Of those, ADAM10 and 17 cleave APP but, to put APP in perspective, ADAM10 and 17 substrates also include a formidable list of type 1 transmembrane proteins that extends beyond Notch, the VEGF and transferrin receptors, the EGF and TNF families, and the large Ig superfamily of cell adhesion molecules (CAMs).
In his Eibsee talk, Saftig noted that deleting ADAM10 and 17 proved to be embryonically fatal and that mouse strains carrying subtler conditional knockouts are in the works. Meanwhile, Saftig and postdoc Karina Reiss have studied cell lines of the original knockouts to assess their ability to still perform shedding, and in this way get a sense of what ADAM10 and 17 do. The overarching finding so far from this ongoing work is that ADAM10/17 cleavage releases two protein stubs, each of which can have a different functional consequence on either side of the membrane.
Saftig focused on cell adhesion molecules (CAMs), a large group of proteins that originally became known for their role in allowing cells to stick together in forming tissues. They also participate in wound healing, inflammation, and tumor metastasis when the balance of cell adhesion versus detachment gets out of kilter. Saftig cited three examples to illustrate principles of ADAM10/17 function. First, consider N-cadherin. It is expressed in fibroblasts but also brain, where it co-localizes with β-catenin and plays a role in synaptic function. The scientists found that ADAM10 releases N-cadherin’s ectodomain, regulating cell adhesion, while internally, this same cleavage alters β-catenin signaling and expression of its target genes (Reiss et al., 2005). Second, ADAM10 turned out to shed E-cadherin in skin cells, and to influence their behavior in this way. Specifically, ADAM10 appears to promote wound healing as a physiological function in skin but, on the flip side, can promote eczema when proinflammatory cytokines induce its expression and excessive E-cadherin shedding then loosens cell-cell contacts in the skin (Maretzky et al., 2005). Third, the cell adhesion protein L1 is known to be important, in development and adult life, for axonal outgrowth and neuronal migration, but it is also present in neuroblastoma. L1 turned out to be constitutively “shedded” by ADAM10, and particular conditions induced additional shedding by ADAM17. The ectodomain locally loosened adhesion and stimulated migration, neurite outgrowth, and synaptic plasticity. L1’s remaining membrane stub became a substrate for none other than γ-secretase (Maretzky et al., 2005).
With APP, ADAM10 shedding also precedes γ-secretase action, but since ADAM10 splits the Aβ sequence in two, subsequent γ-secretase degradation does not release this peptide. The ectodomain of APP, too, is quite potent. Called APPsα, its neuroprotective and growth-promoting properties were worked out a decade ago. This raised the prospect of up-regulating APP shedding as an alternative therapeutic strategy that would kill two birds with one stone by dampening Aβ production while exploiting APPsα’s benefits. Alas, scientists did not know which enzyme did the job in adult human brain. In 1999, Falk Fahrenholz and colleagues at the University of Mainz, Germany, pinpointed ADAM10, at least in human cell lines (Lammich et al., 1999). Later research by Fahrenholz and Rolf Postina, using Fred Van Leuven’s APPV717I transgenic mice, revived interest in this cleavage when it proved the principle that tuning up ADAM10 expression even modestly could prevent amyloid deposition and a learning deficit in the Morris water maze (Postina et al., 2004).
The Fahrenholz group is now exploring ways of cranking up ADAM10 with an eye toward future therapeutics. The Alzforum has previously described ongoing research on the neuropeptide PACAP (see commentary and related news link at Kojro et al., 2006). At the Eibsee, Fahrenholz noted that his group is now attempting to deliver PACAP nasally to mice. A microarray study measuring the effect of chronic ADAM10 up-regulation in the APP-transgenic mice on gene expression found that mRNAs for inflammatory proteins were down, that typical AD-related mRNAs such as BACE and PS were unchanged, and that mRNAs encoding retinoid acid (RA) receptors were more abundant. Fahrenholz pursued this last clue, and this metabolite of vitamin A has since become a focus in the lab. The promoter of the ADAM10 gene contains binding sites for the retinoic acid receptor (Prinzen et al., 2005). Follow-up studies in neuroblastoma cells showed that RA selectively activates expression of ADAM10 but not ADAM17, together with the expression of ADAM10 substrates APP and APLP2. This common up-regulation of enzyme and substrates by retinoic acid might provide enhanced specificity of ADAM10 for APP and APLP2 cleavage (Endres et al., 2005).
RA is not a stranger to AD research. Dietary, metabolic, genetic, and epidemiological studies have implicated it before (see ARF conference report; Puchades et al., 2003; Rinaldi et al., 2003). Adult rats fed a vitamin A-deficient diet develop AD-like pathology (Corcoran et al., 2004), and a hippocampal LTP deficit induced by vitamin A deficiency was reported to be reversed by RA application (Misner et al., 2001). Taken together, the data suggested a working hypothesis whereby vitamin A, through a metabolite, might activate RA-responsive elements and in this way activate ADAM10 to boost the α-secretase pathway of APP processing. To test this concept, the Fahrenholz group is currently keeping mice on a vitamin A-deficient diet and reconstituting it with RA.—Gabrielle Strobel.
Generally speaking, drug development research improves when a range of proteins becomes known that interacts with the target of choice in biologically important ways. A number of labs are now on the trail of proteins that control α-secretase cleavage of APP. While in its early days, this work is already showing that α-secretase may be subject to more intricate regulation than γ-secretase, noted Haass. Stefan Lichtenthaler, at Ludwig-Maximilians University, Munich, described some of those regulators, which he found in an expression cloning strategy devised to identify proteins that activate APP ectodomain shedding (Schoebel et al., 2006). The proteins fell into three groups. Some were intracellular signaling proteins, but their effect was small. A much stronger boost for APP shedding was measured with proteins that reduce endocytosis of APP from the cell membrane (where most α cleavage tends to occur), probably because endocytosis brings APP cheek to jowl with BACE lodged in endosomes. Intriguing proteins in this category include dynamin and a new sorting protein, Snx31. A third group of proteins that regulate APP shedding are intracellular adaptors that mediate the association of APP with other membrane proteins, famously LRP (see below). Broadly speaking, this work dovetails with a hypothesis about abnormalities in endocytosis, autophagy, and ensuing intraneuronal degradation blockages in early AD pathogenesis that Randy Nixon and Anne Cataldo have developed in recent years (see, e.g., Grbovic et al., 2003; Yu et al., 2005).
The endocytosis inhibitors Lichtenthaler found include some known and some novel ones. Known ones are endophilin A3, a member of the endophilin family of trafficking proteins, which activated α-secretase strongly and specifically. Another one is the GTPase dynamin. It mediates endocytosis by way of clathrin-coated pits and is necessary for synaptic vesicle recycling. In 2002, Paul Coleman’s group identified dynamin as a protein whose mRNA plummeted in microarray studies of single neurons from AD brain (see ARF related news story; Yao et al., 2003). Other scientists have since implicated dynamin in synaptic failure in AD (e.g., Kelly et al., 2005), or as a factor influencing APP processing (Chyung and Selkoe, 2003), and a dynamin mutant in Haass’ lab serves as a tool to hold back APP at the plasma membrane.
Dynamin draws added intrigue from a novel protein Lichtenthaler identified. Called Snx31, it belongs to a poorly studied group of sorting nexins (SNXs) that mediate endocytosis in various cell types. Snx31 is highly expressed in brain and strongly stimulates APP shedding, Lichtenthaler reported. Its mechanism is still unclear, but tinkering with its levels yielded phenotypes that mirrored those generated by dynamin manipulation. Possibly, Snx31 and dynamin might reduce APP endocytosis as binding partners, Lichtenthaler speculated. Snx31 appears more specific than merely being another protein in a generalized endocytosis machinery, however. Experiments in C. elegans, done with Ralf Baumeister, suggest that Snx31 is required for endocytosis of only a certain subset of cell surface proteins. Besides APP, an important one appears to be the insulin receptor, raising the possibility that Snx31-mediated endocytosis forms a trafficking nexus connecting APP α cleavage and insulin signaling.
Other APP shedding activators also interfere with endocytosis, though in a different way. APP’s cousin APLP1 came up in the expression screen; its effect just appeared in print (Neumann et al., 2006; see Dominic Walsh’s comment there). In brief, the scientists propose that APLP1 can form a tripartite complex with the transmembrane protein Low Density Lipoprotein Receptor Related Protein (LRP). The adaptor protein Fe65 holds this complex together just below the membrane. Prior work by Claus Pietrzik and others had shown essentially the same thing for APP (Kinoshita et al., 2001; Pietrzik et al., 2004). According to this model, APLP1 would draw LRP molecules away from APP, leaving more APP molecules in a state that is less likely to be endocytosed, and fewer in the LRP complex that gets readily internalized. Fe65 can “glue” together either APP and LRP, or APLP1 and LRP, by binding to a NPXY internalization motif that APP, APLP1, and LRP each carry in their cytoplasmic tails. (Each Fe65 molecule can bind this motif twice.) The rates by which these complexes form, then, may help determine the rate of endocytosis and of APP shedding, Lichtenthaler suggested.
Pietrzik, who is at the University of Mainz, Germany, expanded the LRP story. At 600 kD, LRP is a massive receptor protein, and boasts at least 30 ligands. It is intimately intertwined with AD, beginning with a genetic association and continuing through its role as a receptor for ApoE and α2 macroglobulin. LRP’s manifold interactions with molecules relevant to AD pathogenesis are only beginning to be exposed, but it is already clear that its importance extends beyond its role in lipoprotein metabolism to include strong influences on APP processing. Over the past few years, Pietrzik first developed the model of how LRP, Fe65, and APP might bind together to influence APP internalization and, by extension, APP shedding and Aβ secretion. (Pietrzik et al., 2002). While this model is controversial, Pietrzik pointed to supporting in-vivo data by scientists at Aventis, who bred Fe65-transgenic and APP-transgenic mice and found that Fe65 reduced Aβ levels in the double-transgenics (Santiard-Baron et al., 2005). This fits into Pietrzik’s model in that adding surplus Fe65 to the mix would lead to some Fe65 molecules to bind to LRP and other Fe65 molecules to bind to APP. Yet these dimeric complexes would preclude formation of the trimeric ones that are necessary for APP endocytosis. The result would be more APP shedding even though Fe65’s primary role is to mediate internalization, which ordinarily reduces shedding. This ties into the notion that endocytic processing is a major source of Aβ, an idea long held by Eddie Koo, Gunnar Gouras, and others.
Other work by this group is testing the idea that LRP might influence APP processing in yet another way, that is, by holding APP in early secretory compartments such as the ER, where γ-secretase will not process it. Ongoing work with an ER retention signal on LRP is suggesting that LRP and APP travel together through the secretory pathway, and that this journey may occur in the same tripartite setup with Fe65, Pietrzik said. Their relationship is even more intimate than that, as recent studies have shown that LRP itself first sheds its ectodomain and then undergoes γ-secretase cleavage, complete with LRP-CTF release. In this process, LRP competes with APP (Lleo et al., 2005; Zerbinatti and Bu, 2005). A take-home message from this evolving data is that any researcher studying APP processing would do well to include LRP in the experimental design, as this type 1 membrane protein influences APP at many levels, Pietrzik said.—Gabrielle Strobel.
Interactions of APP with its family members APLP1 and APLP2 have complicated not only the study of its internalization and processing but also the search for its physiological function. That this is not fully understood is frequently cited as one of the gaping holes of AD research. A number of functions have been established, though their precise molecular mechanisms remain elusive. At the Eibsee, Ulrike Mueller, now at the University of Heidelberg, offered an update on her longstanding project to dissect APP function genetically with a series of knockout and, most recently, knock-in mice strains. Mueller made the first APP knockout strain (Mueller et al., 1994), which retained some residual APP expression, as well as a second, complete knockout (Li et al., 1996). The mice’s subtle phenotype initially flummoxed scientists, as they had expected a more dramatic role for this ubiquitous and conserved protein. The mice were small and weak, had small brains with mild anatomic defects, were prone to seizures, and performed poorly in behavioral paradigms of learning and exploration. Mueller subsequently generated various combinations of knockouts and crosses with APLP1 and 2 strains, and in this way found out that these family members were compensating for the physiological roles of APP (Heber et al., 2000; also von Koch et al., 1997). APLP1 and 2 are similar to APP, are both expressed in brain but lack the Aβ sequence. Their compensation made defining precisely what APP itself does a difficult task. To remove all possible interference, Mueller’s group recently generated a triple-knockout strain (Herms, 2004). These mice showed a patterning defect in their cortical architecture that resembled a human disease called type 2 lissencephaly, in which newly born neurons in the developing cortex do not build up their layered architecture in the proper order. The APP/APLP1/APLP2 knockouts also had too few reelin-producing cells.
The question then arose as to which sections of the APP protein mediate which of its functions. To address it, Mueller knocked two different fragments of APP back into the mice lacking APP, and asked which snippets of APP could rescue which aspect of the original APP single-knockout phenotype. This work is ongoing, but initial data suggest that the product of APP shedding by α secretase, i.e., the secreted APPα ectodomain, is able to restore most of the functions lost in the APP single-knockout phenotype. (Results for knock-in rescue of the double-knockout phenotypes are not available yet.) This finding appears to raise new questions about the importance of intracellular AICD signaling and subsequent gene expression to APP function.
The role of the APP cytoplasmic tail in APP function is very much in flux these days, and it came up on a related issue, as well. APP clearly plays a role in the cortical positioning of embryonic neuroblasts, but exactly which parts of it mediate this process is unclear. Tracy Young in Selkoe’s laboratory used in-utero electroporation of RNAi to delete APP from the developing forebrain of mouse embryos. This acute, localized knockdown of APP prevented neuroblasts from migrating up toward the brain’s surface after their birth. Rescue experiments with different parts of APP suggested that its C-terminus is required for cortical migration, while some of Mueller’s studies point to the ectodomain. The phenotypes are similar but not the same. While both manipulations of APP—triple-knockout or acute RNAi—disturbed the migration of cortical neuroblasts, Mueller’s triple-knockout led some to overshoot their proper place in the cortical architecture. Indeed, in certain spots they burst through the pial membrane at the top of the brain, possibly because they failed to stick to the extracellular matrix there. By contrast, Young and Selkoe’s experiment prevented the neuroblasts from ever reaching the cortical layers in the first place; they instead massed near their birthplace at the subventricular zone. The experiments are sufficiently different to preclude direct comparison at this early stage, but the scientists agreed that these apparent discrepancies are important and will be worked out experimentally.—Gabrielle Strobel.
Cell culture studies notwithstanding, in the brain, the presynaptic terminals of neurons are thought to be a main source of Aβ, and a controversy is simmering in the field about exactly how it gets there. One view, advanced by Larry Goldstein at University of California, San Diego, proposes that some is generated in transport vesicles traveling down the axon to the nerve terminal. In a nutshell, this research suggests that APP anchors cargo vesicles to the motor protein kinesin, either directly or via linker proteins in a complex, and that one and the same vesicle tends to contain all necessary components of APP γ cleavage. Axonal transport blockages would drive up Aβ generation because the components would dwell together in the vesicles longer (see Stokin and Goldstein, 2006; Stokin et al., 2005; Kamal et al., 2001; Alzforum axonal transport discussion). A collaboration of scientists from several groups has been unable to reproduce some of the data, leaving the issue open (see Lazarov et al., 2005; Zheng comment there, and Goldstein reply).
At the Eibsee, two speakers reported that they tested Goldstein’s hypothesis with various cell-based approaches but found no support for it. Stefan Kins, at Heidelberg University, Germany, was a coauthor of Lazarov et al., 2005, and has since run further experiments. In the paper, the authors had suggested that a reported interaction between APP’s cytoplasmic tail and the light chain of kinesin might be nonspecific. At the conference, Kins added that he searched for APP/kinesin complexes with coimmunoprecipitation of mouse brain lysates to examine the idea that APP might interact indirectly with kinesin through a cytoplasmic linker, but detected no such complexes. These approaches still leave open the possibility of an indirect association between APP and kinesin mediated through scaffolding proteins such as JIP. JIP1 has been suggested to couple APP-containing vesicles to kinesin motors (see, e.g., Matsuda et al., 2003), and could conceivably do so through its interaction with the NPTY internalization motif on APP’s cytoplasmic tail. To test this directly, Kins used siRNA to knock down JIP1 in cultures of cortical primary neurons, and found that the neurons transported most of their APP from the ER out to neurites regardless of how much JIP1 they had available. JIP might link a small pool of APP to kinesin, but for the bulk of APP the kinesin linker remains elusive, Kins said. Its linker need not even interact with its cytoplasmic tail, as an APP mutant lacking that piece still traveled out to neurites unperturbed.
Finally, Kins noted that APP, APLP1, and APLP2 are transported to different sections of the cell membrane. APLP1 resides primarily in dendritic, postsynaptic membranes, and APLP2 in presynaptic areas and growth cones, where each forms dimers with APP (Soba et al., 2006). For this reason, Kins suggested that each family member may travel in its own type of vesicle, using a different linker protein to a different subset of kinesin motor, and that this helps direct their specific route of transport.
Claire Goldsbury, who works with Eva-Maria Mandelkow at the Max-Planck Unit for Structural Molecular Biology in Hamburg, approached the issue from a different angle. She is interested in the relationship between tau and APP in axonal transport. Goldsbury tested the proposal that APP/BACE/γ-secretase travel in the same vesicles and are cleaved en route. First, she asked whether blocking APP transport in cultured neurons would increase Aβ production. She cultured rat primary neurons, transfected them with a Swedish APP-YFP construct, imaged the neurons, and measured how much Aβ they contained and secreted. As expected based on prior work from Mandelkow’s group, Goldsbury saw that overexpressing tau gummed up the microtubules and blocked the transport of APP from the nucleus out to the neuron’s tips. As planned, this lengthened the time APP vesicles spent in transit, yet Aβ levels never increased, indicating the APP does not get cleaved in transit. This was shown directly by double-labeling APP at both ends with YFP and CFP and watching the ratio of colors during vesicle movement. The ration did not change, indicating that APP molecules remained intact (Goldsbury et al., Traffic, 2006, in press).
Next, Goldsbury reported that vesicles containing APP and BACE move quite differently in culture. APP vesicles move swiftly down the neuron’s processes, whereas BACE vesicles tended to “dawdle,” moving forward a bit, then back, and taking lots of stationary breaks in between. Cotransfection of both APP (labeled yellow) and BACE (labeled red) showed little overlap in vesicles. At the Keystone conference earlier this year, Mandelkow already reported that APP and BACE reside in different vesicles (see ARF conference story).—Gabrielle Strobel.
News on Tau: More Mice Enter the Picture, Structure Takes Shape
The Mandelkow group has been working to generate inducible, TET-off tau-transgenic mice to extend their body of cell-based findings into in-vivo data. At the meeting, Magda Mocanu presented an initial analysis. The group had generated four strains, each expressing endogenous mouse tau plus a modest level (0.5 to 2 times endogenous tau) of a different transgene in their forebrain. The set consists of a full-length and an “abbreviated” version of two types of construct. The abbreviated version involves expression of only the repeat domain of the transgene in question. The transgenes are
a ΔK280 deletion mutation that aggregates aggressively and causes a clinical tauopathy (Rosso et al., 2003) that is, the “proaggregation” strain; and
an “antiaggregation” control obtained by replacing two residues in tau’s most aggregation-prone hexapeptide motifs with prolines, which prevents formation of β-structure and aggregation (von Bergen et al., 2001).
The researchers first asked how aggregation influences pathology. In the case of the full-length proaggregation form of tau, the scientists saw a pathological conformation of tau (i.e., MC1 staining) and hyperphosphorylation at many sites in the hippocampus after 6 months, but no tangle formation. They saw only minor changes in phosphorylation with the antiaggregation mutant. All changes were reversible when tau expression was switched off for 6 weeks. In the case of the short form of tau, the proaggregation mutant began developing tangles in the entorhinal cortex at 3 months and progressed with age, even at low levels of exogenous tau (0.7 of endogenous), whereas the antiaggregation mutants showed no tangles. The proaggregation short form of tau nucleated aggregates, which then grew to incorporate both transgenic and endogenous tau. In old mice, the proaggregation, but not the antiaggregation mutants caused synaptic loss and neurodegeneration in the dentate gyrus of the hippocampus. In addition, this tau showed the redistribution from the axonal to the somatodendritic compartment that is characteristic of AD, and it showed enhanced phosphorylation at the KXGS motifs that control microtubule interactions. Switching off the short proaggregation form of tau for 6 weeks after 10 months of expression reversed the mislocalization and phosphorylation of soluble tau, but the aggregated protein remained largely in place. A direct comparison with other tau-transgenic mouse strains, including those made by the Davies, Ashe, and LaFerla labs, can be made once analysis of these mice is more advanced.
Fred Van Leuven of the University of Leuven, Belgium, presented new data in support of his view that tangles may not be the primary offender in tauopathy. Instead, Van Leuven said, the true causes of neurodegeneration—while still elusive—appear prior to, or in parallel to, the tangles. A similar shift is well underway in the amyloid field (Moechars et al., 1999; Dewachter et al., 2002 and many other investigators), but is only beginning in the tau field. For one, Van Leuven presented data on a mouse line that expresses both the tau kinase GSK3β and the human tauopathy mutation P301L. These double-transgenics display a heavy load of neurofibrillary tangles in their hippocampus and cortex, showing them in practically every neuron visible in microscopic sections. Yet despite this massive pathology, the mice survive normally, probably because GSK3β overexpression somehow lessened tauopathy in the mice’s hindbrain and spinal cord. (Mice transgenic for P301L alone do die early, see Terwel et al., 2004; the authors believe that the tauopathy in hindbrain kills them.) Besides emphasizing that regional differences between different brain areas contribute to what ends up being pathogenic, these data suggest that tangles per se do not necessarily cause neurodegeneration, said Van Leuven. Other factors, some having to do with a predeposition interplay between Aβ and tau, do the neurons in, and they still need to be discovered.
Along this line of thought, Van Leuven crossed APP-V717 mice, which have intraneuronal amyloid, diffuse and neuritic plaques, and vascular amyloid, with P301L mice that show intraneuronal tau filaments and die at 1 year of age. Initial analysis of this new, combined model indicates that amyloid pathology is more extensive, and tau pathology dramatically more intense in the forebrain than in the respective single-transgenic lines. Curiously, though, this combined model, too, has less tauopathy in the hindbrain, than the P301L single-transgenics and the doubly afflicted mice live longer, too. Confusing as this is, said Van Leuven, these two studies have in common that they both point to GSK3β as a factor linking Aβ and tau pathogenesis. This connection has been made by studies of lithium and of Aβ oligomer immunization (see, e.g., Ma et al., 2006).
Finally, Van Leuven added new data to the study of p25/Cdk5. Prior work by Li-Huei Tsai, Van Leuven, and others has put this kinase on the AD map, and LaFerla’s group has linked fibrillar Aβ to inflammation and tau hyperphosphorylation via p25/Cdk5 (Kitazawa et al., 2005). At the Eibsee, Van Leuven added data by showing that inducible p25 transgenic mice suffer severe neurodegeneration with drastic loss of brain volume that is mediated not primarily by tauopathy but by early microgliosis and astrogliosis. In yet another demonstration that transgenic mice can be unpredictable and bewildering, Tsai’s inducible p25 transgenic mice do show tau pathology (Cruz et al., 2003). Rather than being a model for tauopathy, Van Leuven views his new mouse as a window into hippocampal sclerosis, a common cause of adult epilepsy (see Sen et al., 2006).
Frank LaFerla reviewed recently published research on his group’s ongoing analysis of the triple-transgenic mouse model (e.g., Oddo et al, 2006; Billings et al., 2005). He emphasized that, overall, this study to date suggests that Aβ facilitates subsequent tau pathology by three pathways.
One, fibrillar Aβ tends to act via inflammatory processes in that amyloid induces cytokines that, in turn, activate the tau kinase Cdk5 (Kitazawa et al., 2005).
Second, LaFerla believes that Aβ oligomers interfere with the proteasome’s ability to clear tau (Oddo et al., 2004). On this issue, Haass questioned how intraneuronal Aβ would interact with the proteasome from its primary site of residence within the lumen of endosomes and late endosomes (also called multivesicular bodies). LaFerla noted that the triple-transgenic mice have large quantities of Aβ across different intraneuronal organelles, even some in the cytosol. A study published last month by Gunnar Gouras’s lab may shed light on this issue (see Almeida et al., 2006). In neurons from human AD brain, Aβ was spotted in multivesicular bodies (Takahashi et al., 2002).
Third, various tau kinases, including GSK3β, become more active in response to rising Aβ levels (Oddo et al., 2006).
Theoretically, axonal transport effects might constitute a fourth link, but data on that question is not available yet.
Importantly, the phosphorylation state of tau, more than its absolute levels, determines whether tau pathology can reverse after Aβ removal. Stimulation of the cholinergic system with M1 agonists also affects both pathologies (Caccamo et al., 2006). This approach will see its definitive test in the clinic as TorreyPines Pharmaceuticals tests NXG267; other pharmaceutical companies are said to take a second look at such agonists, as well.
Clearly, compensatory processes wrestle with toxic ones at the early stages of tau pathology, and their outcome determines the cell’s fate. One path toward understanding how tau goes bad early on is to understand its structure, a line of inquiry spearheaded by Eckhard Mandelkow. At the conference, Mandelkow started by noting that the repeat domain of tau is central to its aggregation. Imaging tau is notoriously difficult because tau is natively unfolded. It is highly soluble, and probably resists folding because it contains unusually few hydrophobic amino acids for a protein of its size. Indeed, its partially aggregated oligomeric forms clearly come up in Mocanu’s transgenic mice prior to paired helical filaments, but even they are unstable and too floppy to be viewed in the EM, as has been done for other amyloid oligomers, let alone be crystallized for x-ray imaging.
How to nail this pudding to the wall, then? The Mandelkow group has used other structural approaches to get a feeling for tau’s conformations. Tau contains two hexapeptide motifs flanking the repeat 2 domain that form stable β structures and seed tau aggregation. The ΔK280, as well as the P301L mutation responsible for FTDP-17, make precisely those areas more hydrophobic and prone to assume a β-structure (von Bergen et al, 2001; von Bergen et al., 2005). Mandelkow calls these motifs VQIINK and VQIVYK (pronounced VIKING and VIKIVIK, respectively). Their study received an independent boost last year when David Eisenberg at the University of California, Los Angeles, published their crystal structure (Nelson et al., 2005). It was taken from a prion protein and the paper related primarily to the structural conversion of cellular prion protein to its pathogenic alter ego, yet a similar motif occurs in Aβ and in tau (see also Inouye et al., 2006). Eisenberg proposed a broader concept of an “amyloid spine,” whereby in all these amyloid building blocks the hydrophobic side chains of the seeding motif face each other to form a tight surface that interacts through van der Waals forces. Called a steric zipper, it is based in part on the principle of protein folding that hydrophobic residues tend to turn inside, in this case in the form of two β strands leaning against each other. Eisenberg’s framework fits with Mandelkow’s own model of these kernels of nascent β-structure, as well as with his group’s simultaneous description of the tau repeat domain by NMR solution structure (Mukrasch et al., 2005, done in collaboration with Christian Griesinger’s group at the Max-Planck Institute for Biophysical Chemistry in Goettingen).
Another way of making educated guesses about a protein’s structure is FRET, a method where scientists attach pairs of fluorophores along a protein’s length and then measure fluorescence when these pairs come close together as the protein folds. With this approach, Mandelkow’s group recently found that tau in solution does not actually exist as a totally random coil as one might expect for natively unfolded proteins. Instead, it assumes a sort of paper clip structure that, while still flexible, nevertheless shows the C-terminus bending over to lie in the middle between the repeat domain and the N-terminus, close enough to give FRET with both (Jeganathan et al., 2006; see image below).
It’s still unclear how this paper clip structure relates to early AD pathology, but the widely used tau antibodies MC1 and Alz50 (Jicha et al., 1997) provide an intriguing clue, Mandelkow said. They are thought to pick up a misfolded and pathogenic, but pretangle, conformation of tau; they, too, recognize a composite epitope made up of amino acids from both the N-terminal and the repeat domain of tau.—Gabrielle Strobel.
Therapy development efforts in AD have diversified to include statins, PPARγ agonists, antioxidants, and certain NSAIDs as existing drug candidates, as well as the tau kinase GSK3β as a target for preclinical drug development. But most of the effort is still centered around the amyloid hypothesis. At the conference, two speakers detailed their efforts to find a better NSAID.
Sascha Weggen told the audience what his lab has done since he set up shop at the University of Mainz. By studying NSAIDs, Weggen can test whether Aβ42 truly is a good target in AD. This stands to reason because many FAD mutations increase the levels of this peptide only modestly (see ARF Discussion). Conceivably, Weggen said, a modest increase in the Aβ42/Aβ40 ratio over many years might lead to sporadic AD, and an equally modest reduction might therefore stave off disease. Initial presenilin inhibitors have proven toxic, but NSAIDS offer a new chance to tackle the question.
NSAIDs became a hot topic in AD research when epidemiological studies detected protection against AD in chronic users, but figuring out how they might do so has been all but straightforward. These mechanisms have been proposed:
Cox inhibition, (few AD researchers believe it plays the major role)
PPARγ activation to affect expression of inflammatory mediators (e.g., IL-1b, IL-6)
PPARγ activation to reduce BACE expression
γ-secretase modulation to lower Aβ42
the Rho-Rock pathway (several groups have largely ruled this out)
inhibition of Aβ aggregation
reduced expression of α1-antichymotrypsin, an acute phase protein that accelerates amyloid pathology in APP-transgenic mice (this has only been shown for ibuprofen)
sAPPα stimulation
Some of these ideas rest exclusively on in-vitro data, and Weggen offered some negative data of the kind that rarely see formal publication. When his group used pulse-chase experiments to test a claim for stimulation of sAPPα production and a subsequent signaling pathway (Avramovich et al., 2002), the scientists were unable to reproduce these findings in the same cells as originally reported. Seeing no effect with ibuprofen and indomethacin on sAPPα secretion, Weggen concluded that sAPPα stimulation is not an alternative mechanism of NSAID action.
Weggen focuses on the fourth category, that is, NSAIDS that lower Aβ42 production while leaving Aβ40 unchanged and increasing Aβ38 production. They include sulindac sulfide, ibuprofen, indomethacin, and flurbiprofen. New experiments into their mechanism of action show that they are unlikely to cause the mechanism-based toxicity that bedeviled previous γ-secretase inhibitors (Weggen et al., 2001; Lanz et al., 2005). Even so, their COX activity has gastrointestinal side effects that preclude their chronic, much less preventive, use at high doses. The way forward, then, would lie with new compounds that modulate γ-secretase in this desired way but don’t block COX.
Aβ-lowering NSAIDs do not modulate γ-secretase through the active site. They probably tweak a third, allosteric binding site proposed for γ-secretase, and several labs are in hot pursuit of exactly where they interact with members of the complex. For his part, Weggen noticed that certain FAD mutations change the secretase’s response to the NSAIDs. For example, wild-type cells produce less Aβ42 when treated with sulindac sulfide, but cells carrying certain FAD mutations that aggressively increase Aβ42 production do not. This suggests that the residues changed by these FAD mutations, or conformational effects of these mutations, affect the NSAID binding site.
Clinical studies of existing NSAIDs have largely failed or been halted, though an ongoing phase 3 trial of a purified stereoisomer may yet be successful (see R-flurbiprofen story). While encouraging, even this compound does not fill the need for new modulators that are more potent, penetrate the brain better, and stay active longer. To this end, Weggen has teamed up with Herbert Waldman at the Max-Planck Institute in Dortmund, Germany, whose collection of 500 sulindac sulfide and indomethacin analogs Weggen is screening for Aβ42-lowering and against COX activity. The scientists will test the hits in long-term animal models. Incidentally, Weggen noted, this will allow him to test whether anti-inflammatory effects have contributed to the AD protection reported in the epidemiological NSAID studies, after all. This possibility has gotten lost amid the current focus on Aβ42 lowering, but inflammation has never been formally ruled out.
Like Weggen, Boris Schmidt, who is at the Technical University of Darmstadt, Germany, searches for Aβ42-lowering compounds that are more potent than the four drugs known to do so. These four have IC50s that are in the micromolar range, and that is too high for wide use. Schmidt, a medicinal chemist, tested all commercially available NSAIDs in Germany for their ability to change the Aβ42/40/38 ratio and made about 150 analogs to selected ones, but none had the desired properties. The most potent one proved to be carprofen, a prescription drug approved for use in dogs only. (The only legal way to obtain this drug from pharmacies, even for scientific study in cells, is to obtain an arthritic dog, Schmidt said, adding, tongue in cheek, that his grant application for such an animal is pending.)
Schmidt also picked up research by Greg Cole and Sally Frautschy on Aβ lowering by the curry spice curcumin, and found that curcumin is an all-out inhibitor of γ-secretase but would be toxic at the concentrations needed. His lab made milder analogs that fit the desired γ-secretase modulation profile in the low micromolar range. Schmidt’s group also analyzed the mechanism of action of many γ-secretase modulators reported in the literature.
As take-home lessons of these studies, Schmidt noted that modulation is fundamentally different from inhibition, as the former need not change total Aβ load much at all. Secondly, Schmidt found that all modulators that achieve a favorable ratio of Aβ42/40/38 have a carboxylic residue in common and that certain lipophilic substitutions can improve the compounds’ potency.
Schmidt’s other effort has been to use NSAIDs as “bait” to “fish” for individual γ-secretase complex components in order to define the NSAID binding site. This fishing expedition has not caught anything whatsoever, Schmidt said, and the lesson to be learned is that the true binding site of the NSAID γ-secretase modulators most likely comprises a protein-protein interaction that gets disturbed by the bait and its attached label.
Overall, Schmidt said, the field of AD drug discovery is advancing but as of yet, no brain-penetrating β- or γ-secretase inhibitors have made it into a phase 3 trial. In Schmidt’s estimation, too, allosteric modulation may be a better ticket to success.
No matter which position they take on the amyloid hypothesis, scientists agree that its definitive test will occur in the clinic. Christoph Hock of the University of Zurich, offered 3-year follow-up data on the Zurich cohort of the aborted AN1792 active immunotherapy trials. (To read up on this topic, see Fox et al., 2005; Gilman et al., 2005; Orgogozo et al., 2003; Hock et al., 2003; Alzforum discussion; workshop coverage on AN1972 biomarker use.) Hock showed a movie sequence of the first Zurich patient receiving a shot of the AN1792 vaccine into his upper left arm. A second film sequence showed a different, 68-year-old trial participant taking a line-marking test that is part of the trial’s cognitive assessment package. This man had developed the meningoencephalitis (ME) with an MRI lesion, a deficit in this particular test, but few other symptoms. He recovered from the encephalitis, and his mild-to-moderate AD still remains stable, Hock said. By contrast, another trial patient who developed ME had a normal MRI throughout, even though he was severely ill. “This just shows how difficult these trials are,” Hock said. He recalled a challenging time in 2002, as the investigators found themselves under assault from newspapers and colleagues who accused them of rushing into human tests prematurely. Since then, Hock and colleagues have observed the patients in hopes of learning if the stalled trial can still yield information about the underlying questions of whether immunotherapy can remove amyloid and whether that changes the disease.
In short, the trial appears to have worked inasmuch as it induced antibodies that crossed the blood-brain barrier and removed patches of parenchymal amyloid. People who developed high levels of antibody tended to show an improvement on certain memory tests, Hock said. In the overall AN1792 cohort, the control group declined less steeply than the control group of the Zurich cohort did, so the total measured effect came out small. Among the Zurich patients, the responders still decline more slowly on MMSE than the non-responders after 3 years, Hock noted. An additional MRI scan at 36 months showed that hippocampal volume in the responders is still at the level measured at 24 months. At 24 months, it had begun to recover after an unexpected decline at 12 months. “This is an indication that there might be a restoration of hippocampal volume after the immunization,” Hock said. Three years after the immunization, antibody titers, hippocampal volume, and cognitive performance correlated, indicating that the immune response is long-lasting. At 30, the number of patients in this cohort was small to begin with. Now, close to 4 years after the trial began, about 20 percent are dropping out of the follow-up study every year, though the six highest responders are still doing well, Hock said.
Despite its setbacks, the AN1792 trial has stimulated interest in immunotherapy to where these companies are now said to pursue their own programs: Elan and Wyeth; Novartis; Hoffman LaRoche; Eli Lilly and Co.; Rinat Neuroscience; Neurochem; Eisei Co./BioArctic Neuroscience; Abmaxis Inc./BioArctic; AC Immune; AC-Immune (Lausanne); Boehringer Ingelheim; Merck/Acumen; and Abbot, with all except Elan/Wyeth at the preclinical or phase 1 stage.—Gabrielle Strobel.
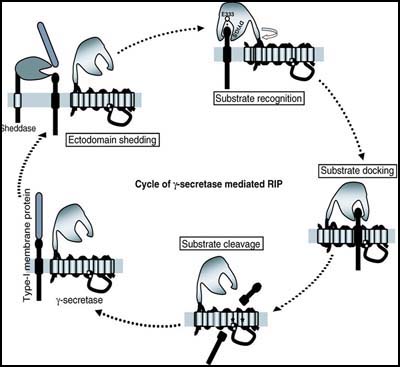


Comments
No Available Comments
Make a Comment
To make a comment you must login or register.