IFITM3 Forges Link Between Neuroinflammation and Aβ Production
Quick Links
Neuroinflammation and Aβ deposition are two hallmarks of AD, and now, a study published September 3 in Nature is forging an alluring mechanistic link between them. Researchers led by Yueming Li at Memorial Sloan Kettering Cancer Center in New York reported that interferon-induced transmembrane protein 3 (IFITM3) binds to γ-secretase, revving up the enzyme’s production of Aβ peptides. Knocking down IFITM3 squelched Aβ production in human cells and in a mouse model of amyloidosis. In the human brain, levels of the protein rose with age and in people with AD, and correlated with the amount of inflammatory cytokines and viral proteins present in the brain, as well. In all, the findings link age-related neuroinflammation with increased production of Aβ, and highlight IFITM3 as a potential therapeutic target.
- IFITM3 binds γ-secretase, ramping up Aβ production.
- Knocking out IFITM3 reduces Aβ deposition in 5xFAD mice.
- In human brain, IFITM3 increases with age and in AD.
“The paper is very convincing and springs a new surprise in the study of the γ-secretases,” wrote Bart De Strooper of KU Leuven in Belgium. “It actually turns the classical view that inflammation is a consequence of amyloid plaque accumulation upside-down, providing mechanistic support for the hypothesis that inflammation causes increased Aβ generation.”
Beyond its four essential subunits of presenilin, nicastrin, Aph1, and Pen-2, the γ-secretase complex hosts some hangers-on that modulate its activity in different ways. Researchers are keen to find ways to selectively block the enzyme’s processing of amyloid-β precursor protein (APP), while sparing the cleavage of other important substrates such as Notch. To that end, scientists have developed small molecule γ-secretase modulators (GSMs) that, besides having clinical potential, are able to identify proteins that associate with active γ-secretase complexes.
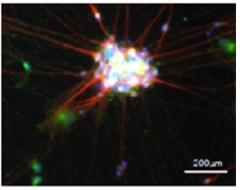
IFITM3 Shines. IFITM3 (green) colocalizes (white) with the neuronal protein MAP2 (red) in this human induced pluripotent stem cell-derived neuron. [Courtesy of Hur et al., Nature, 2020.]
By attaching a photo-affinity probe to GSMs and then flashing them with UV light to trigger cross-linking with adjacent proteins, researchers hope to capture proteins likely to play a physiological role in the enzyme’s function. While at Merck, Li used this approach, albeit with γ-secretase inhibitors, to help peg PS1 as the catalytic subunit of γ-secretase (Li et al., 2000; Xu et al., 2002).
For the current study, co-first authors Ji-Yeun Hur and Georgia Frost used a photo-affinity labeled version of E2012—a GSM created by Eisai but no longer in clinical development—to fish out endogenous protein modulators of γ-secretase (Apr 2011 conference news; Aug 2013 news). Even when conjugated with the photoaffinity probe BPyne, E2012 binds to PS1 N terminal fragments within active γ-secretase complexes (Pozdnyakov et al., 2013). The researchers mixed neuronal cell membranes with E2012-BPyne, induced cross-linking with UV light, purified the BPyne-labeled complexes, and identified two proteins via western blot: the N-terminal fragment of PS1, as predicted, and IFITM3.
An interferon-inducible protein involved in antiviral responses, IFITM3 had previously been spotted among PS1’s interactors in a proteomics screen led by De Strooper (Oct 2009 news on Wakabayashi et al., 2009). Li’s group used several complementary approaches, including co-immunoprecipitation with various PS1 antibodies, to confirm that indeed, IFITM3 bound to PS1 within active γ-secretase complexes in mouse primary neurons. IFITM3 also interacted with PS2.
This interferon-induced protein is expressed in many cell types, including mouse hippocampal neurons, astrocytes, and microglia. Human induced pluripotent stem cell-derived neurons and primary human astrocytes make the protein (see image above). In cells lacking both PS1 and PS2, IFITM3 protein levels plunged, suggesting that presenilins stabilize it.
Does IFITM3 influence γ-secretase activity? To investigate, the researchers knocked down IFITM3 in human embryonic kidney (HEK) cells. Without it, production of Aβ40 and Aβ42 peptides dropped by 17 and 24 percent, respectively. In an astrocytoma cell line lacking IFITM3, the amyloid peptides dropped by 36 and 27 percent, respectively. The opposite was true for γ-secretase cleavage products of Notch, which ramped up without IFITM3. The findings suggest that IFITM3 enhances processing of APP, but suppresses processing of Notch.
In the brains of wild-type mice, IFITM3 levels rose with age, more than doubling between the ages of 4 and 28 months. What’s more, IFITM3 associated with γ-secretase only in the older mice. The researchers generated IFITM3 knockout mice, and found that γ-secretase production of Aβ40 and Aβ42 dropped by 15 and 24 percent, respectively, in brain tissue extracted from 18-month-old knockouts relative to wild-type.
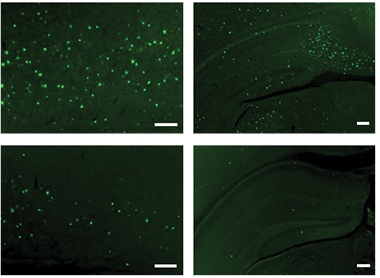
Plaque Protection. Fewer amyloid plaques (green) arise in the cortex (left) and hippocampus (right) of 5xFAD when they lack IFITM3 (bottom). [Courtesy of Hur et al., Nature, 2020.]
IFITM3 levels also ramped up with age in the 5xFAD mouse model of amyloidosis. At 12 months of age, IFITM3 levels in 5xFAD mice were nearly double those in wild-type. These transgenic mice overexpress human APP and PS1, which likely also influences IFITM3 expression levels. While IFITM3 primarily localized to blood vessels and meninges in wild-type mice, in plaque-ridden 5xFAD mice the protein also comingled with astrocytes and microglia. The researchers crossed 5xFAD mice to IFITM3 knockouts, and found that loss of IFITM3 not only reduced the production of Aβ peptides by γ-secretase, but also dramatically lessened Aβ plaque burden. Compared with 5xFAD controls, 5xFAD lacking IFITM3 had 54 and 81 percent fewer plaques in the hippocampus and cortex, respectively, at 4 months (see image above).
What about in people? The researchers assessed IFITM3 expression in postmortem brain samples from multiple cohorts. In the Genotype-Tissue Expression (GTEX) cohort, they measured more IFITM3 transcripts in the hippocampus and cortex with age: IFITM3 expression approximately doubled between the ages of 20 and 70. In a Mayo Clinic cohort, IFITM3 transcripts were more abundant in the temporal cortices of 80 people with AD than in 76 controls (see figure below). In a separate cohort of frontal cortex samples from the UCSD AD Research Center (ADRC), 18 people with AD had more IFITM3 mRNA and protein than did 10 controls.

Ticking Up in AD. In the Mayo Clinic cohort, IFITM3 transcripts in the temporal cortex (left) were more abundant in people with late-onset AD (LOAD) than in controls. The same was true in the frontal cortex (right). [Courtesy of Hur et al., Nature, 2020.]
Although IFITM3 protein was more abundant on average, there was high variability in IFITM3 among people with LOAD in the UCSD cohort. To see if IFITM3 levels correlated with γ-secretase activity, Hur and colleagues divided the samples into high and low IFITM3 bins, then measured APP processing in cell membranes extracted from each. Lo and behold, the eight LOAD samples with high IFITM3 protein cranked out 127 and 130 percent more Aβ40 and Aβ42, respectively, than did controls, or LOAD samples with low IFITM3. The findings suggest that IFITM3 correlates with higher Aβ production. In support of this idea, IFITM3 expression levels correlated with Aβ load in multiple brain regions among postmortem samples from the Mount Sinai Brain Bank Dataset.
What dictates IFITM3 expression in the brain? Treating cell cultures with either type I or type II interferons boosted IFITM3 expression and ramped up production of Aβ. Dovetailing with this, IFITM3 expression correlated with pro-inflammatory cytokine levels in brain samples from Harvard Brain Tissue Resource Center, and with herpesvirus proteins and hepatitis C infection in the Mount Sinai brain samples. Together, the findings suggest that inflammatory stimuli—such as viral infections or age-related cellular stress—might enhance IFITM3 expression, leading to stepped-up production of Aβ.
The findings jibe with the notion that Aβ peptides are rallied to contain microbes (May 2016 news; Jun 2018 news). IFITM3 has been implicated in defense against numerous viruses, including influenza, Zika, and most recently SARS-CoV-2.
“This work directly links Aβ production with innate immunity and neuroinflammation and provides a novel mechanism as to how Aβ secretion is stimulated in response to an invading pathogen,” wrote Huaxi Xu, co-editor in chief at Molecular Neurodegeneration. “This not only provides insight into Aβ’s function as an antimicrobial peptide, but also establishes IFITM3 as a potential therapeutic target to reduce Aβ production.”
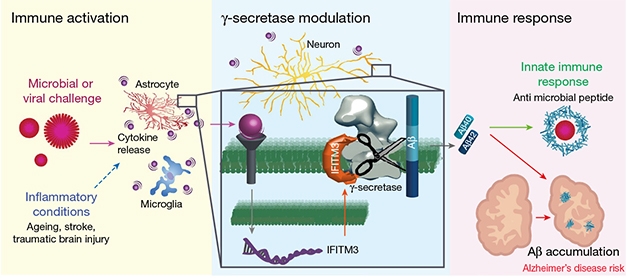
Feel the Burn. Glial cells secrete pro-inflammatory cytokines in response to inflammatory stimuli such as viral infections (left). This enhances IFITM3 expression in neurons and other cells, upping Aβ (middle). Aβ peptides corral microbes, and can accumulate, leading to AD (right). [Courtesy of Hur et al., Nature, 2020.]
“Taken together, this broad and deep study suggests that IFITM3 may be a safe target for the treatment of AD, lowering Aβ production without inhibiting Notch signaling, the latter a well-known liability of γ-secretase inhibitors,” commented Michael Wolfe of the University of Kansas in Lawrence.
Li told Alzforum that efforts are already underway to develop IFITM3 inhibitors. In a comment to Alzforum, Weiming Xia of Boston University School of Medicine agreed that this might be a promising therapeutic approach. “While efforts have been made to explore other γ-secretase interacting proteins as therapeutic targets for AD in the past two decades, IFITM3 will likely outperform those previous targets for a number of reasons, mainly anti-inflammation/Aβ dual efficacies, existing potent compounds as candidate IFITM3 inhibitors, and anti-aging potential,” he wrote. Tamping down IFITM3 expression with brain-penetrant, anti-inflammatory drugs is one potential strategy, Xia noted. Another is to modify existing small molecule GSMs to thwart IFITM3’s binding to PS1.
“The identification of a subpopulation of LOAD patients in whom IFITM3 expression strongly correlates with γ-secretase activity suggests that IFITM3 may be used as a biomarker to stratify AD patients,” added Xu. “As LOAD is a multifactorial disease, identification of biomarkers for subpopulations of AD is invaluable to studying underlying mechanisms and developing targeted therapeutics.”—Jessica Shugart
References
News Citations
- Barcelona: Allosteric γ Modulation Moves Toward Clinic
- More Evidence that γ-Secretase Modulators Spare Essential Substrates
- New Spin on γ-Secretase—Tangled in Tetraspanin Web?
- Like a Tiny Spider-Man, Aβ May Fight Infection by Cocooning Microbes
- Herpes Triggers Amyloid—Could This Virus Fuel Alzheimer’s?
Paper Citations
- Li YM, Xu M, Lai MT, Huang Q, Castro JL, DiMuzio-Mower J, Harrison T, Lellis C, Nadin A, Neduvelil JG, Register RB, Sardana MK, Shearman MS, Smith AL, Shi XP, Yin KC, Shafer JA, Gardell SJ. Photoactivated gamma-secretase inhibitors directed to the active site covalently label presenilin 1. Nature. 2000 Jun 8;405(6787):689-94. PubMed.
- Xu M, Lai MT, Huang Q, DiMuzio-Mower J, Castro JL, Harrison T, Nadin A, Neduvelil JG, Shearman MS, Shafer JA, Gardell SJ, Li YM. gamma-Secretase: characterization and implication for Alzheimer disease therapy. Neurobiol Aging. 2002 Nov-Dec;23(6):1023-30. PubMed.
- Pozdnyakov N, Murrey HE, Crump CJ, Pettersson M, Ballard TE, Am Ende CW, Ahn K, Li YM, Bales KR, Johnson DS. γ-Secretase Modulator (GSM) Photoaffinity Probes Reveal Distinct Allosteric Binding Sites on Presenilin. J Biol Chem. 2013 Apr 5;288(14):9710-20. PubMed.
- Wakabayashi T, Craessaerts K, Bammens L, Bentahir M, Borgions F, Herdewijn P, Staes A, Timmerman E, Vandekerckhove J, Rubinstein E, Boucheix C, Gevaert K, De Strooper B. Analysis of the gamma-secretase interactome and validation of its association with tetraspanin-enriched microdomains. Nat Cell Biol. 2009 Nov;11(11):1340-6. PubMed.
Further Reading
Primary Papers
- Hur JY, Frost GR, Wu X, Crump C, Pan SJ, Wong E, Barros M, Li T, Nie P, Zhai Y, Wang JC, Tcw J, Guo L, McKenzie A, Ming C, Zhou X, Wang M, Sagi Y, Renton AE, Esposito BT, Kim Y, Sadleir KR, Trinh I, Rissman RA, Vassar R, Zhang B, Johnson DS, Masliah E, Greengard P, Goate A, Li YM. The innate immunity protein IFITM3 modulates γ-secretase in Alzheimer's disease. Nature. 2020 Oct;586(7831):735-740. Epub 2020 Sep 2 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
UK Dementia Research Institute@UCL and VIB@KuLeuven
This is really nice work from Drs. Hur, Frost, and a team of collaborators led by Yue-Ming Li. Yue-Ming belongs to a small group of researchers who did pioneering work on the γ-secretase complex (or better yet, complexes, each having very different biological functions). In the current work, they are building further on their observations with various types of γ-secretase inhibitors and modulators to identify a novel, very interesting regulator of γ-secretase activity, IFITM3, which enhances APP processing (and lowers Notch processing) in cell cultures.
It is nice to see that this protein was also identified in our own work many years ago where we tried to identify regulators and interactors of γ-secretase via a proteome approach (Wakabayashi et al., 2009). They also provide good initial correlative evidence to show human relevance of the role of IFITM3 in Aβ generation and potentially in Alzheimer’s disease.
The paper is very convincing and springs a new surprise in the study of the γ-secretases. It actually turns the classical view that inflammation is a consequence of amyloid plaque accumulation upside-down, providing a mechanistic support for the hypothesis that inflammation causes increased Aβ generation. Yue-Ming and colleagues discuss the different implications from their observations very nicely in their manuscript, so no need to repeat them here.
Overall, the story links together, in a very interesting way, two major trains of thought in the Alzheimer’s field—Aβ generation and inflammation. The γ-secretases are pivotal in a vicious cycle that encompasses both and provides different entry points for environmental factors and aging to initiate the Alzheimer’s disease-causing cascade.
References:
Wakabayashi T, Craessaerts K, Bammens L, Bentahir M, Borgions F, Herdewijn P, Staes A, Timmerman E, Vandekerckhove J, Rubinstein E, Boucheix C, Gevaert K, De Strooper B. Analysis of the gamma-secretase interactome and validation of its association with tetraspanin-enriched microdomains. Nat Cell Biol. 2009 Nov;11(11):1340-6. PubMed.
Boston University School of Medicine
This work on a γ-secretase-modulating protein IFITM3 published by Yue-Ming Li's group is a sequel to his Nature article on presenilin-labeling active site γ-secretase inhibitors published in 2000. The γ-secretase complex and its modulators have been extensively studied in the past 20 years (Xia, 2019). A number of γ-secretase-interacting proteins have been identified, including CD147, p23/TMP21, γ-secretase-activating protein (GSAP), as well as Hif-1α by the Li group.
IFITM3, a member of the interferon-induced transmembrane protein family reported in the current Nature article, is different from previously identified γ-secretase interacting proteins. While efforts have been made to explore GSAP and other γ-secretase-interacting proteins as therapeutic targets for Alzheimer's disease (AD) in the past two decades, IFITM3 will likely outperform those previous targets for a number of reasons, mainly anti-inflammation/Aβ dual efficacies, existing potent compounds as candidate IFITM3 inhibitors, and anti-aging potentials.
First, agents targeting IFITM3 probably carry similar properties of a previous γ-secretase-modulating compound that entered clinical trial, R-flurbiprofen. This nonsteroidal anti-inflammatory drug (NSAID) inhibits interferon γ (IFNγ) and selectively reduces the 42-residue of amyloid-β protein (Aβ42). However, in clinical trials, R-flurbiprofen did not achieve statistical significance on either of its primary endpoints—cognition or activities of daily living. R-flurbiprofen is a weak γ-secretase modulator (GSM) with an IC50 for Aβ reduction of approximately 300 μM. Due to its poor brain penetration, it was unlikely to have lowered brain Aβ42 levels in the clinical studies. In the future, brain-permeable potent IFITM3 inhibitors, once identified, should carry desirable efficacy against neuroinflammation akin to NSAIDs, and modulate γ-secretase activity to reduce Aβ42 production in the central nervous system.
Second, we have a battery of potent γ-secretase modulators as candidate IFITM3 inhibitors. IFITM3 was identified through its binding to a well-characterized γ-secretase modulator E2012. E2012 is a non-NSAID-derived compound that inhibits both Aβ40 and Aβ42 production, consistent with Li's Nature report that suppressing IFITM3 exhibits a similar reduction of Aβ40 and Aβ42. Another non-NSAID derivative GSM is the former NeuroGenetics’ Compound 4, which directly interacts with presenilin and PEN-2 and inhibits all Aβ peptide production (Aβ40 and 42) in animals under chronic treatment. Other GSMs with IC50 at sub-μM include GSM1, GSM-10h, EVP-A, EVP-B, JNJ-40418677, and more potent BMS-932481 and BMS-986133 (with IC50s for reducing Aβ42 of 6.6 and 3.5 nM, respectively ), as well as BPN-15606 (IC50 of 7 nM and 17 nM to reduce Aβ42 and Aβ40, respectively). Efforts by medicinal chemists could be devoted to modifying existing GSMs, exploring their inhibitory effects on IFITM3, and establishing their structure-activity relationships.
Third, inhibiting IFITM3 will likely suppress Aβ and other aging-related pathological processes. Since aging upregulates IFITM3 and is the primary risk factor for AD, targeting IFITM3 clearly presents a unique opportunity to potentially modulate cellular processes, especially those related to neuroinflammation and cognitive function. While we continue to explore the mechanisms of action of anti-IFITM3 agents, prevention of memory loss during aging is a key efficacy readout for any clinical trial of AD therapeutics.
References:
Xia W. γ-Secretase and its modulators: Twenty years and beyond. Neurosci Lett. 2019 May 14;701:162-169. Epub 2019 Feb 11 PubMed.
University of Kansas
In the more than 20 years of study of γ-secretase and its role in AD, many high-profile reports have touted the discovery of modulatory proteins with potential as therapeutic targets, only leading to a disappointing inability to validate and extend those findings. Despite this frustrating history, this new report by Hur et al., that interferon-induced transmembrane protein 3 (IFITM3) is a modulator of γ-secretase activity, is exciting, with important implications for AD pathogenesis, diagnostics, and treatment. Independent validation and extension are still essential, but the thorough and rigorous nature of this study and the large multi-institutional team of investigators bodes well for the future.
In human cell lines and primary neurons, IFITM3 associated with the γ-secretase complex to increase its proteolytic processing of amyloid-β precursor protein to Aβ40 and Aβ42: Knockdown or knockout of IFITM3 reduced this activity, while overexpression increased it. In contrast, IFITM3 deficiency increased γ-secretase cleavage of Notch1, revealing promising substrate-selective modulatory effects. Photoreactive small molecule probes, derived from a γ-secretase Aβ42-lowering modulator (E2012) as well as an active site-directed inhibitor (L-685,458) covalently labeled the Presenilin-1 (PSEN1) N-terminal fragment (NTF) subunit of γ-secretase as well as IFITM3, suggesting these two proteins are proximal. This was confirmed with an L-685,458 analog containing two photoreactive groups, which was capable of crosslinking PSEN1 NTF to either the PSEN1 CTF subunit or to IFITM3, demonstrating that IFITM3 is in proximity to the protease active site.
IFITM3 expression increased with age in mice, as did its association with γ-secretase. Knockout of IFITM3 did not affect levels of γ-secretase components but did lower Aβ40 and Aβ42 production by the protease and substantially decreased amyloid plaque deposition in the 5XFAD mouse model. Provocatively, IFITM3 expression was increased in late-onset AD human brain, and γ-secretase production of Aβ40 and Aβ42 was significantly increased in a subset of brain samples with the highest IFITM3 expression. Moreover, treatment with interferons could stimulate IFITM3 expression and increase γ-secretase-dependent production of Aβ40 and Aβ42 in cultured neurons and astrocytes, and IFITM3 expression in human brains positively correlated with expression of a variety of cytokines and with expression of two human viruses (a herpes virus and a hepatitis C virus).
Taken together, this broad and deep study suggests that IFITM3 may be a safe target for the treatment of AD, lowering Aβ production without inhibiting Notch signaling, the latter a well-known liability of γ-secretase inhibitors. IFITM3 also has potential as a diagnostic marker for late-onset AD, with high levels of expression possibly a strong risk factor. Perhaps most exciting is the potential mechanistic link between AD and neuroinflammation, the underappreciated third pathological feature of AD after amyloid plaques and tau tangles. Inflammatory states that increase cytokine levels in the brain may also increase γ-secretase-mediated production of Aβ.
Finally, the connection between viral expression and IFITM3 provides further evidence that increased Aβ production may be a response to microbial infection. While Aβ may help in the fight against microbes, the long-term consequences could be increased risk of AD.
Molecular Neurodegeneration
The involvement of the immune system in Alzheimer’s disease (AD) is well-established. Immune activity is thought to play several roles in the progression of AD pathology, which may be both beneficial and detrimental, notably: neuroinflammation and microglial clearance of amyloid-β plaques (Blasko and Grubeck-Loebenstein, 2003; Shi and Holtzman, 2018).
Furthermore, recent work has indicated that the Aβ peptide, which is considered a primary cause of AD, functions as an antimicrobial peptide (AMP), and therefore, plays a role in the innate immune system. In both human and animal models, infection leads to increased amyloid-β production in the brain and Aβ oligomers can form fibrils that entrap pathogens and damage cell membranes (Kumar et al., 2016; Eimer et al., 2018). This suggests that Aβ may be produced as a defense against invading pathogens. How Aβ production is coordinated in such an immune response remains an outstanding question in the field. In addition, the consequence of pathogen-stimulated Aβ production on risk of developing AD has yet to be investigated.
In this Nature paper, Hur et al. identify a novel mechanism by which an immune response can stimulate Aβ production, providing a link between defense against infection and AD pathology. The authors used the γ-secretase modulator (GSM) E2012-BPyne, which interacts with presenilin 1 (PS1), to search for novel GSM binding proteins, and identified interferon-induced transmembrane protein 3 (IFITM3). IFITM3 is an interferon-stimulated protein known to restrict various viral infections (Bailey et al., 2014). The authors determined that IFITM3 directly binds to γ-secretase near the active site and reduces the production of Aβ40 and Aβ42.
Notably, when IFITM3-/-mice were crossed to 5xFAD transgenic mice, there was a striking reduction in the number of Aβ plaques in the cortex and hippocampus. Furthermore, IFITM3 expression increases with age, the biggest risk factor for AD. Analyzing postmortem tissue, Hur et al. identified a subpopulation of human patients with late-onset Alzheimer’s disease (LOAD) who had elevated IFITM3 levels. Tissues with high IFITM3 also had significantly increased γ-secretase activity for Aβ40 and Aβ42 production, further highlighting that IFITM3 functions as a γ-secretase modulatory protein.
This work directly links Aβ production with innate immunity and neuroinflammation and provides a novel mechanism by which Aβ secretion is stimulated in response to an invading pathogen. This not only provides insight into Aβ’s function as an AMP, but also establishes IFITM3 as a potential therapeutic target to reduce Aβ production. The identification of a subpopulation of LOAD patients in whom IFITM3 expression strongly correlates with γ-secretase activity suggests that IFITM3 may be used as a biomarker to stratify AD patients. As LOAD is a multifactorial disease, identification of biomarkers for subpopulations of AD is invaluable to studying underlying mechanisms and developing targeted therapeutics.
References:
Blasko I, Grubeck-Loebenstein B. Role of the immune system in the pathogenesis, prevention and treatment of Alzheimer's disease. Drugs Aging. 2003;20(2):101-13. PubMed.
Shi Y, Holtzman DM. Interplay between innate immunity and Alzheimer disease: APOE and TREM2 in the spotlight. Nat Rev Immunol. 2018 Dec;18(12):759-772. PubMed.
Kumar DK, Choi SH, Washicosky KJ, Eimer WA, Tucker S, Ghofrani J, Lefkowitz A, McColl G, Goldstein LE, Tanzi RE, Moir RD. Amyloid-β peptide protects against microbial infection in mouse and worm models of Alzheimer's disease. Sci Transl Med. 2016 May 25;8(340):340ra72. PubMed.
Eimer WA, Vijaya Kumar DK, Navalpur Shanmugam NK, Rodriguez AS, Mitchell T, Washicosky KJ, György B, Breakefield XO, Tanzi RE, Moir RD. Alzheimer's Disease-Associated β-Amyloid Is Rapidly Seeded by Herpesviridae to Protect against Brain Infection. Neuron. 2018 Jul 11;99(1):56-63.e3. PubMed.
Bailey CC, Zhong G, Huang IC, Farzan M. IFITM-Family Proteins: The Cell's First Line of Antiviral Defense. Annu Rev Virol. 2014 Nov 1;1:261-283. PubMed.
Vrije Universiteit
GGZinGeest/Amsterdam UMC
This study provides new information about a molecular pathogenic mechanism that linked innate immunity activation to increased Aβ production by an interferon-induced transmembrane protein that upregulates γ-secretase activity. Seeing the genetic evidence for the involvement of the innate immunity in the etiology of late-onset AD, these findings raise the question whether interferons, as interferon-γ, are related to the genetic background of late-onset AD.
Twin studies have shown that the cytokine production capacity is under strong genetic control (Craen et al., 2005). We have studied the cytokine production capacity in ex vivo stimulated full blood samples from middle-aged offspring with and without a parental history of late-onset AD (van Exel et al., 2009). We found that the production capacity of some proinflammatory proteins, including interferon-γ, was significantly higher in offspring with a parental history of AD upon stimulation of whole blood with LPS. These findings were independent of ApoE4 genotype. This study provides evidence for a proinflammatory genotype for late-onset AD that is characterized by higher production capacity for proinflammatory cytokines including interferon-γ.
It seems from a neuroinflammatory perspective that AD has two faces (Eikelenboom and van Gool, 2004). In one type such as the familial autosomal-dominant form of AD, the neuroinflammatory response follows the increased production and deposition of Aβ. In a second type, the late-onset and most common form of AD, a neuroinflammatory response precedes the process of increased of Aβ generation.
References:
de Craen AJ, Posthuma D, Remarque EJ, van den Biggelaar AH, Westendorp RG, Boomsma DI. Heritability estimates of innate immunity: an extended twin study. Genes Immun. 2005 Mar;6(2):167-70. PubMed.
van Exel E, Eikelenboom P, Comijs H, Frölich M, Smit JH, Stek ML, Scheltens P, Eefsting JE, Westendorp RG. Vascular factors and markers of inflammation in offspring with a parental history of late-onset Alzheimer disease. Arch Gen Psychiatry. 2009 Nov;66(11):1263-70. PubMed.
Eikelenboom P, Van Gool WA. Neuroinflammatory perspectives on the two faces of Alzheimer's disease. J Neural Transm. 2004 Mar;111(3):281-94. PubMed.
University of Texas Health Science Center at Houston
Baylor College of Medicine
We read the study by Hur et al. from Dr. Yue-ming Li’s group with great interest and are impressed with their insightful discoveries on IFITM3.
Previously, we uncovered a type I interferon response in brains harboring amyloid plaques from both mouse models and human AD (Roy et al., 2020). We showed that certain amyloid plaques are capable of stimulating microglia to mount an intrinsic antiviral immune response, which upregulates a panel of interferon-stimulated genes (ISGs) including IFITM3. Moreover, interferon activates complement cascade and induces synapse loss. Given the ability of IFITM3 to further ramp up Aβ production and plaque formation, AD is seemingly afflicted by a pathogenic feed-forward loop involving APP processing, innate immune activation, and action of distinct ISGs.
In Aβ models, early and persistent interferon and antiviral immune response was recently confirmed by an analysis of microglial proteomes (Monasor et al., 2020). We have detected IFITM3 as an ISG expressed by plaque-associated microglia, which display features of microglial neurodegenerative phenotype (Krasemann et al., 2017). Not only was IFITM3 upregulated in activated mouse microglia, IFITM3 mRNA levels were also elevated in the brain tissues of dementia patients archived in Mount Sinai Brain Bank (Roy et al., 2020). Further, we detected IFITM3 protein expression in microglia enveloping Aβ plaques in human AD brain.
Despite being recognized as a protein involved in antiviral protection, the precise role played by microglial IFITM3 in the context of AD is not known at this time. Given the fresh interest in therapeutically targeting IFITM3, it would be pertinent to evaluate the drug candidate’s effect on microglia and other cell types in conjunction with that on γ-secretase from neurons and astrocytes.
References:
Krasemann S, Madore C, Cialic R, Baufeld C, Calcagno N, El Fatimy R, Beckers L, O'Loughlin E, Xu Y, Fanek Z, Greco DJ, Smith ST, Tweet G, Humulock Z, Zrzavy T, Conde-Sanroman P, Gacias M, Weng Z, Chen H, Tjon E, Mazaheri F, Hartmann K, Madi A, Ulrich JD, Glatzel M, Worthmann A, Heeren J, Budnik B, Lemere C, Ikezu T, Heppner FL, Litvak V, Holtzman DM, Lassmann H, Weiner HL, Ochando J, Haass C, Butovsky O. The TREM2-APOE Pathway Drives the Transcriptional Phenotype of Dysfunctional Microglia in Neurodegenerative Diseases. Immunity. 2017 Sep 19;47(3):566-581.e9. PubMed.
Sebastian Monasor L, Müller SA, Colombo AV, Tanrioever G, König J, Roth S, Liesz A, Berghofer A, Piechotta A, Prestel M, Saito T, Saido TC, Herms J, Willem M, Haass C, Lichtenthaler SF, Tahirovic S. Fibrillar Aβ triggers microglial proteome alterations and dysfunction in Alzheimer mouse models. Elife. 2020 Jun 8;9 PubMed.
Roy ER, Wang B, Wan YW, Chiu G, Cole A, Yin Z, Propson NE, Xu Y, Jankowsky JL, Liu Z, Lee VM, Trojanowski JQ, Ginsberg SD, Butovsky O, Zheng H, Cao W. Type I interferon response drives neuroinflammation and synapse loss in Alzheimer disease. J Clin Invest. 2020 Apr 1;130(4):1912-1930. PubMed.
Make a Comment
To make a comment you must login or register.