New Myelin Makes Memories, but Supply Wanes with Age
Quick Links
While the vast majority of myelin sheaths in the brain are laid down early in life, a pair of new studies published February 10 in Nature Neuroscience reveal that a fresh supply of the fatty axonal conductor is required to establish and maintain memories in the adult brain. One, led by Jonah Chan and Mazen Kheirbek at the University of California, San Francisco, reported that newly minted, myelin-producing oligodendrocytes cemented unpleasant memories in mice. The other, led by Chan and Feng Mei at Third Military Medical University in Chongqing, China, reported that the birth of new oligodendrocytes plummets with age—a slowdown that could underlie age-related memory loss. In both, a drug that fosters the growth of new oligodendrocytes improved memory.
- During memory formation, oligodendrocytes mature and make myelin.
- These new myelin-producing cells are required for long-term memory.
- Oligodendrocyte differentiation wanes with age; restoring it prevents memory loss.
The studies add to growing evidence that active myelination plays a crucial role in memory function, and mesh with recent studies implicating myelination malfunctions in neurodegenerative disease, according to Chan.
Michael Lipton of Albert Einstein College of Medicine in New York agreed. “Even more provocative is the evidence that remyelination can restore function,” he wrote. “Dependent upon confirmation that the same mechanisms are operative in humans, this result points toward potential interventions, either behavioral or pharmacologic, that can promote remyelination and ultimately might restore lost function.”
Part and parcel of most functional axons, myelin sheaths speed up the conductance of neuronal signals in the brain. While much of the myelin in the brain is as old as the axons themselves, a fraction of myelin continues to be produced by a small pool of new oligodendrocytes that develop throughout life, in response to new experiences and learning. This experience-dependent myelination is thought to bestow structural plasticity on the brain (for review, see Mount and Monje, 2017).
Such myelination has been implicated in motor learning, and drugs that interfere with oligodendrocyte function reportedly cause memory deficits in mice (Xiao et al., 2016; May 2019 news). In addition, recent studies implicate myelin degeneration in neurodegenerative diseases associated with memory loss, including AD and vascular dementia (Benitez et al., 2013; Gao et al., 2011; Nasrabady et al., 2018).
What is myelination’s role in memory? Chan and Kierbek addressed this fundamental question in their study, which probed relationships between oligodendrocytes, myelin, and fear memory in mice. First author Simon Pan and colleagues used a contextual fear-conditioning model of memory, in which mice receive a series of foot shocks in a specific context. When placed back into that context at a later time, the animals freeze, signifying recall of the prior zaps. The researchers found that indeed, the conditioned mice froze when placed back into the shock context 24 hours and 30 days later, suggesting both recent and remote memories had been formed.
The researchers found that within 24 hours of fear conditioning, oligodendrocyte progenitor cells (OPCs) started multiplying in the prefrontal cortex. Using a mouse model in which only freshly matured oligodendrocytes become fluorescent, the researchers tracked differentiation of the cells over time. They found that the OPCs differentiated into oligodendrocytes rapidly, and that these cells gradually matured into myelin-producing oligodendrocytes over the next month.

Myelin Makes Memories? Seven days after fear conditioning (left), OPCs had differentiated into new oligodendrocytes (green) that did not yet produce myelin (purple). By 30 days (right), the new oligodendrocytes had matured into myelin-producing cells (white). [Courtesy of Pan et al., Nature Neuroscience, 2020.]
Were these fresh oligodendrocytes supporting memory formation? To find out, the researchers used mice in which Myrf, a transcription factor necessary for oligodendrocyte differentiation, can be switched off in OPCs. When the researchers shut down the birth of newborn oligodendrocytes in mice and then subjected them to fear conditioning, the animals recognized the context of the foot shock 24 hours after receiving it. However, they froze less often than control mice when placed into the shock context 30 days later, suggesting that production of new myelin-producing cells was essential for remote, but not recent, memories.
Stopping oligodendrocyte differentiation also squelched stereotypical neuronal responses that happen when memories are made and recalled. Shortly after mice were put back into the environment in which they had received a foot shock a month earlier, neurons in brain regions associated with fear ramped up expression of Fos, an immediate early gene involved in memory recall. This fundamental response was absent in mice that were unable to make new oligodendrocytes. Curiously, the researchers found that treating the mice with clemastine fumarate enhanced remote fear memory and increased Fos expression in fear regions of the brain. Clemastine fumarate is an over-the-counter antihistamine with pro-myelinating activity (Green et al., 2017).
Together, the findings suggest that a learning experience sparks the proliferation and rapid differentiation of OPCs into oligodendrocytes, which mature into myelin-producing cells over the course of several weeks. This fresh supply of myelin somehow influences the reorganization of neuronal circuitry that forms the basis of memory, Chan proposed.
If new oligodendrocytes are important in memory, might a slowdown in their production underlie age-related memory loss? Co-first authors Fei Weng and Shu-Yu Ren, both in Chongqing, and colleagues addressed this question in the second paper. They started by tracking myelin production in mice with age. Using transgenic mice in which newly formed oligodendrocytes and myelin sheaths can be inducibly labeled, the researchers spotted numerous myelin-producing newbies in four- to six-month-old mice in the corpus callosum, but by 13 months of age, new oligodendrocytes were few and far between.
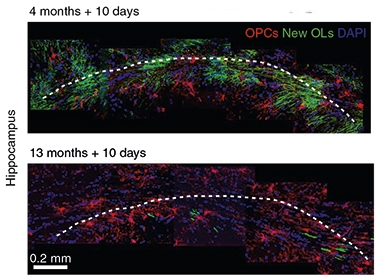
Old Mice, Old Myelin. Ten days after switching on a gene that labels new oligodendrocytes (green), the hippocampi of 4-month-old mice (top) are chock-full of fresh cells, while 13-month-olds have few. [Courtesy of Wang et al., Nature Neuroscience, 2020.]
As the spigot of fresh myelin started to pinch, age-related memory deficits emerged. Compared with 4-month-old mice, 13-month-olds took longer to learn the location of a submerged platform in the Morris water maze test of spatial memory. The hippocampus is crucial for storage of spatial memory, and the researchers found far fewer newly minted, myelin-producing oligodendrocytes in the CA1 region of the hippocampus in 13-month-old than in younger adult mice. Expression of myelin basic protein (MBP)—the building block of myelin sheaths—was also lower in the hippocampi of older mice.
Wielding a menagerie of mouse models, the researchers went on to reveal that blocking differentiation of oligodendrocytes led to memory loss in 4-month-old mice, while revving up the maturation of new oligodendrocytes prevented memory deficits in older mice, and even upped the density of synaptic puncta in the hippocampus. Similarly, treating mice with clemastine prevented the dip in spatial memory with age.
Finding that active myelination plays a crucial role in memory was new and exciting, Chan told Alzforum, but much remains to be understood about the mechanisms involved. How new myelin affects neural networks that form and cement memories is unclear. Also uncertain is whether freshly made myelin can replace myelin lost to degeneration, either in aging or neurodegenerative disease. While myelin loss is the primary event in multiple sclerosis, Chan noted that the role of myelin degeneration in diseases such as AD, where researchers tend to focus on the loss of neurons, is less obvious. “Are neurons dying as a cause or consequence of myelin degeneration?” he asked. Altered oligodendrocyte gene expression has been recently reported in AD brain, implicating the cells in the disease (Mathys et al., 2019; May 2019 conference news).—Jessica Shugart
References
News Citations
- Microglia Instigate ‘Chemofog’ by Squelching Myelination
- When It Comes to Alzheimer’s Disease, Do Human Microglia Even Give a DAM?
Paper Citations
- Mount CW, Monje M. Wrapped to Adapt: Experience-Dependent Myelination. Neuron. 2017 Aug 16;95(4):743-756. PubMed.
- Xiao L, Ohayon D, McKenzie IA, Sinclair-Wilson A, Wright JL, Fudge AD, Emery B, Li H, Richardson WD. Rapid production of new oligodendrocytes is required in the earliest stages of motor-skill learning. Nat Neurosci. 2016 Sep;19(9):1210-1217. Epub 2016 Jul 25 PubMed.
- Benitez A, Fieremans E, Jensen JH, Falangola MF, Tabesh A, Ferris SH, Helpern JA. White matter tract integrity metrics reflect the vulnerability of late-myelinating tracts in Alzheimer's disease. Neuroimage Clin. 2013;4:64-71. Epub 2013 Nov 9 PubMed.
- Gao J, Cheung RT, Lee TM, Chu LW, Chan YS, Mak HK, Zhang JX, Qiu D, Fung G, Cheung C. Possible retrogenesis observed with fiber tracking: an anteroposterior pattern of white matter disintegrity in normal aging and Alzheimer's disease. J Alzheimers Dis. 2011;26(1):47-58. PubMed.
- Nasrabady SE, Rizvi B, Goldman JE, Brickman AM. White matter changes in Alzheimer's disease: a focus on myelin and oligodendrocytes. Acta Neuropathol Commun. 2018 Mar 2;6(1):22. PubMed.
- Green AJ, Gelfand JM, Cree BA, Bevan C, Boscardin WJ, Mei F, Inman J, Arnow S, Devereux M, Abounasr A, Nobuta H, Zhu A, Friessen M, Gerona R, von Büdingen HC, Henry RG, Hauser SL, Chan JR. Clemastine fumarate as a remyelinating therapy for multiple sclerosis (ReBUILD): a randomised, controlled, double-blind, crossover trial. Lancet. 2017 Dec 2;390(10111):2481-2489. Epub 2017 Oct 10 PubMed.
- Mathys H, Davila-Velderrain J, Peng Z, Gao F, Mohammadi S, Young JZ, Menon M, He L, Abdurrob F, Jiang X, Martorell AJ, Ransohoff RM, Hafler BP, Bennett DA, Kellis M, Tsai LH. Single-cell transcriptomic analysis of Alzheimer's disease. Nature. 2019 Jun;570(7761):332-337. Epub 2019 May 1 PubMed.
Further Reading
Papers
- Hase Y, Horsburgh K, Ihara M, Kalaria RN. White matter degeneration in vascular and other ageing-related dementias. J Neurochem. 2018 Mar;144(5):617-633. Epub 2018 Jan 9 PubMed.
Primary Papers
- Pan S, Mayoral SR, Choi HS, Chan JR, Kheirbek MA. Preservation of a remote fear memory requires new myelin formation. Nat Neurosci. 2020 Feb 10; PubMed.
- Wang F, Ren SY, Chen JF, Liu K, Li RX, Li ZF, Hu B, Niu JQ, Xiao L, Chan JR, Mei F. Myelin degeneration and diminished myelin renewal contribute to age-related deficits in memory. Nat Neurosci. 2020 Feb 10; PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.