Divide and Conquer: Structure-Function Victory With Presenilin 1
Quick Links
Creating inhibitors for γ-secretase has felt a bit like trying to slay an unseen beast. That’s because this fearsome protein complex—which slashes a precursor into the Aβ peptides that clog the brains of Alzheimer disease patients—has resisted years of intense effort to unravel its structure. At long last, one piece of the massive γ-secretase has succumbed. Researchers led by Volker Dötsch of Goethe University, Frankfurt, Germany, have zeroed in on the catalytic subunit, presenilin 1 (PS1), and report the structure of its C-terminal fragment in this week’s PNAS Early Edition. "It's a pretty big deal,” Michael Wolfe, Brigham and Women’s Hospital, Boston, told ARF. “This is the first detailed structure of any γ-secretase component.” Applying a complementary approach, a Japanese research team led by Taisuke Tomita and Takeshi Iwatsubo, University of Tokyo, has used biochemical experiments to tease out the function of individual transmembrane domains in PS1. Those findings were published online 23 April in the Journal of Biological Chemistry. Together, the two studies nudge the field closer to the possibility of designing γ-secretase inhibitors in a rational manner.
Previous electron microscopy studies gave tantalizing glimpses of various nooks and crannies (Lazarov et al., 2006; Ogura et al., 2006), and more recent EM work revealed the overall shape of γ-secretase to 12-angstrom resolution (Osenkowski et al., 2009)—still a far cry from the fine detail offered by crystallography. Nevertheless, these EM studies, combined with biochemical approaches, have converged on a fairly consistent picture of PS1’s N-terminal fragment. By and large, this fragment appears to have classical transmembrane topology, with six alpha-helices spanning the length of the membrane. The C-terminal fragment (CTF) of PS1 is comparatively smaller but more complex, inspiring continued debate over how many times it crosses the membrane and what those transmembrane regions look like.
To help resolve the controversy, first author Solmaz Sobhanifar and colleagues performed nuclear magnetic resonance (NMR) microscopy on PS1’s C-terminal fragment (CTF) in detergent-solubilized micelles. They describe in their PNAS paper a structural model suggesting that CTF passes through the membrane three times (see figure below). One transmembrane region (shown in green) is a half helix with the catalytic aspartate 385 poking into PS1’s water-filled cavity, consistent with the idea that γ-secretase cleaves amyloid precursor protein (APP) within an intramembrane hydrophilic pore. The second transmembrane domain (shown in yellow) resembles a classical alpha-helix. The third region does not seem to traverse the membrane so much as dip into it. This element actually consists of two short helical regions (shown in red and orange)—one parallel to the membrane surface, the other perpendicular—with a loop between them. Unexpectedly, the model revealed an additional alpha-helix (shown in blue) outside the membrane in the N-terminal region of CTF. Curiously, this new structural element contains several amino acids that are mutated in familial AD, suggesting it could play a functional role.
NMR Peak-a-Boo
Spectroscopy gives first structural model of presenilin 1’s C-terminal fragment (CTF). Three transmembrane regions are shown—a half-helix (green) containing the active site aspartate, a classical alpha-helix (yellow), and a pair of perpendicular shorter helices (red and orange). In this model, CTF has an additional alpha-helix (blue) located away from the membrane. Image credit: Volker Dötsch
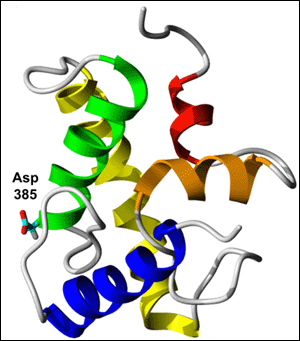
Though an important step toward understanding how γ-secretase acts at an atomic level, the structural data should not be taken “as gospel,” Wolfe cautioned. “Like other structural studies, this provides a model. We can now use the model to generate more specific hypotheses and test them.”
The authors themselves acknowledge that their NMR investigation of highly concentrated, SDS-solubilized CTF raises concern about the biological relevance of these structures. However, given biochemical data hinting that PS1’s cleavage activity happens inside a watery pore (Sato et al., 2006; Tolia et al., 2006), they reasoned that SDS micelles might in fact provide a more natural environment compared to solid membranes that lack other components of the hydrophilic cavity. “They expressed the protein and had to re-dissolve it, and you worry that you wouldn't get homogeneous conformers,” Wolfe told ARF. “But apparently they did, and what they saw by NMR seems largely consistent with other reports using biochemical and molecular approaches, which makes one even more confident in their results.” Furthermore, the authors note that other NMR studies of detergent-solubilized membrane proteins have yielded biologically relevant information.
Given the challenges of probing PS1 through crystallographic or NMR approaches, the Japanese researchers used clever biochemical methods to do structure-function analysis of the protein. As reported in their JBC paper, one strategy entails exchanging individual helices in PS1 with helices of unrelated transmembrane proteins, and then measuring function of the modified PS1s. The researchers have used this transmembrane domain (TMD)-swap approach to show that all six TMDs within the NTF are needed for γ-secretase activity (Watanabe et al., 2005). In the current study—which includes some data initially reported at a 2008 Keystone meeting (see ARF conference story)—first author Naoto Watanabe and colleagues apply the same technique across the entire PS1, including two transmembrane helices in the CTF.
They used the swap method to figure out which transmembrane domains contribute to substrate binding. Prior research reveals an unusual feature of PS1—namely, that initial substrate binding occurs at a site distinct from the enzyme’s active site. Substrates are believed to dock onto PS1 somewhere outside the lipid bilayer before gaining access to the active site in the middle of the membrane, where the cutting takes place.
To determine if a specific transmembrane region is required for PS1 to bind substrate, the scientists used an assay where binding was measured by how well PS1 is labeled with a photoreactive APP helical probe. In this set-up, the probe did not label the TMD2 and TMD6 swap mutants, suggesting that these two PS1 regions are involved in forming the initial substrate-binding site. Furthermore, chemical crosslinking experiments with the photoreactive APP probes revealed that TMD2 and TMD6 are found near TMD9, a region prior research has implicated in substrate binding. Wolfe noted in an email that while TMD2 and TMD6 do appear critical to formation of the initial binding site, whether each region is in fact a part of the docking site is unclear. The uncertainty stems from somewhat weak evidence for TMD2’s proximity to TMD9, he wrote. (See full comment below.)
Taken together, the two papers suggest that a dual-pronged approach may eventually provide the structural information required for rational design of inhibitors to the monstrous and multi-faceted γ-secretase. “Only through a combination of biochemical assays, as in the Watanabe et al. paper, and structural studies, as in ours, might it be possible to design inhibitors that selectively suppress the cutting of the precursor protein without affecting its other targets,” Dötsch wrote (see full comment below).—Esther Landhuis
References
News Citations
Paper Citations
- Lazarov VK, Fraering PC, Ye W, Wolfe MS, Selkoe DJ, Li H. Electron microscopic structure of purified, active gamma-secretase reveals an aqueous intramembrane chamber and two pores. Proc Natl Acad Sci U S A. 2006 May 2;103(18):6889-94. PubMed.
- Ogura T, Mio K, Hayashi I, Miyashita H, Fukuda R, Kopan R, Kodama T, Hamakubo T, Iwatsubo T, Iwastubo T, Tomita T, Sato C. Three-dimensional structure of the gamma-secretase complex. Biochem Biophys Res Commun. 2006 May 5;343(2):525-34. PubMed.
- Osenkowski P, Li H, Ye W, Li D, Aeschbach L, Fraering PC, Wolfe MS, Selkoe DJ. Cryoelectron microscopy structure of purified gamma-secretase at 12 A resolution. J Mol Biol. 2009 Jan 16;385(2):642-52. PubMed.
- Sato C, Morohashi Y, Tomita T, Iwatsubo T. Structure of the catalytic pore of gamma-secretase probed by the accessibility of substituted cysteines. J Neurosci. 2006 Nov 15;26(46):12081-8. PubMed.
- Tolia A, Chávez-Gutiérrez L, De Strooper B. Contribution of presenilin transmembrane domains 6 and 7 to a water-containing cavity in the gamma-secretase complex. J Biol Chem. 2006 Sep 15;281(37):27633-42. PubMed.
- Watanabe N, Tomita T, Sato C, Kitamura T, Morohashi Y, Iwatsubo T. Pen-2 is incorporated into the gamma-secretase complex through binding to transmembrane domain 4 of presenilin 1. J Biol Chem. 2005 Dec 23;280(51):41967-75. PubMed.
Further Reading
Papers
- Tolia A, De Strooper B. Structure and function of gamma-secretase. Semin Cell Dev Biol. 2009 Apr;20(2):211-8. PubMed.
- Osenkowski P, Li H, Ye W, Li D, Aeschbach L, Fraering PC, Wolfe MS, Selkoe DJ. Cryoelectron microscopy structure of purified gamma-secretase at 12 A resolution. J Mol Biol. 2009 Jan 16;385(2):642-52. PubMed.
- Lazarov VK, Fraering PC, Ye W, Wolfe MS, Selkoe DJ, Li H. Electron microscopic structure of purified, active gamma-secretase reveals an aqueous intramembrane chamber and two pores. Proc Natl Acad Sci U S A. 2006 May 2;103(18):6889-94. PubMed.
- Ogura T, Mio K, Hayashi I, Miyashita H, Fukuda R, Kopan R, Kodama T, Hamakubo T, Iwatsubo T, Iwastubo T, Tomita T, Sato C. Three-dimensional structure of the gamma-secretase complex. Biochem Biophys Res Commun. 2006 May 5;343(2):525-34. PubMed.
- Watanabe N, Tomita T, Sato C, Kitamura T, Morohashi Y, Iwatsubo T. Pen-2 is incorporated into the gamma-secretase complex through binding to transmembrane domain 4 of presenilin 1. J Biol Chem. 2005 Dec 23;280(51):41967-75. PubMed.
Primary Papers
- Sobhanifar S, Schneider B, Löhr F, Gottstein D, Ikeya T, Mlynarczyk K, Pulawski W, Ghoshdastider U, Kolinski M, Filipek S, Güntert P, Bernhard F, Dötsch V. Structural investigation of the C-terminal catalytic fragment of presenilin 1. Proc Natl Acad Sci U S A. 2010 May 25;107(21):9644-9. PubMed.
- Watanabe N, Image Image II, Takagi S, Tominaga A, Image Image I, Tomita T, Iwatsubo T. Functional analysis of the transmembrane domains of presenilin 1: participation of transmembrane domains 2 and 6 in the formation of initial substrate-binding site of gamma-secretase. J Biol Chem. 2010 Jun 25;285(26):19738-46. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University of Kansas
This report provides a very thorough and systematic analysis of the role of the various transmembrane domains of PS1 in γ-secretase assembly, stability and activation. The evidence largely supports the role of specific TM domains in the various steps in the maturation of the protease complex as laid out schematically in Figure 9. This includes the role of TM2 and the luminal region of TM6 in the formation of the initial substrate binding site: swapping these regions with those of another unrelated membrane protein leads to the inability of a photoaffinity helical peptide probe targeted to this initial substrate binding site from labeling PS1. These two TM regions are apparently critical to the formation of this binding site, but whether they are both direct contributors to the binding site is not clear. It could be, for instance, that TM2 is not part of the initial substrate binding site but can nevertheless affect the conformation of this binding site. Some chemical crosslinking evidence is provided that TM2 is near TM9, the latter previously being implicated in binding and lateral gating of substrate. However, the evidence for TM2 proximity to TM9 is rather weak, and such proximity would not mean that TM2 is necessarily part of the substrate binding site.…More
View all comments by Michael WolfeThese authors take one of the helices of presenilin at a time, replace it
with a helix of an unrelated protein and then measure its function. If the
helix they exchange is important for a certain function, then the modified
presenilin should no longer be able to show this function. It is like
exchanging the individual parts of a car with, say, parts of household
appliances. If you exchange the motor of the car with a vacuum cleaner and
then try to start the car and it won't, then you have found out that the
motor is necessary to drive the car. However if you push this car it will
still move, so the motor is obviously important for driving but not in
general for the ability of the car to be moved.…More
In essence, Watanabe et al do the same thing with presenilin and its
helices. The n-terminal domain has 6 helices (the C-terminal domain, of
which we have determined the structure has 3). By exchanging every domain
individually, the authors found that helix2 and helix6 are involved in
binding of helical peptides such as βamyloid precursor protein. They
also found that helix9, which is part of our structure, is involved in
substrate binding. Understanding where the substrate binds is very
important. As a field, we need lots of information about which sites bind
specific substrates and which are involved in processing of APP. Only with
such very detailed information will we able to develop drugs that inhibit
formation of the Aβ peptide but not other functions presenilin is
involved in.
In this sense both this and our paper provide complementary information
View all comments by Volker Doetschand provide another small but important piece of the puzzle.
The University of Tokyo
The main finding of this study is that using NMR studies in SDS micelles, the authors determined the structure of PS1 CTF. The structure fits well with previous studies using cysteine accessibility methods by us and by Bart de Strooper’s group. This is important because it is a first structure of a part of PS1 resolved at the atomic level. The study’s potential impact could come from an intriguing feature it shows around catalytic aspartate residue 385, that is, a half helix and extended structure kinked by a glycine residue located near the aspartate. This supports the notion that intramembrane cleavage is occurring within the water-accessible pore structure in the membrane.
At the same time, further study (e.g., using holoprotein) is required, as the PS1 N-terminal fragment (NTF) is also required for the proteolytic activity. Moreover, we have identified that TMD1 of PS1 also faces the catalytic pore (Takagi et al., unpublished result), suggesting that several TMDs in NTFs are involved in the formation of the catalytic pore. …More
Without an atomic structure, it is impossible to “rationally design” inhibitors or modulators. The Doetsch paper is a first step toward understanding the process by γ-secretase at an atomic level. The field has encountered difficulties for structural studies of this unusual membrane-bound protease; however, we also know that tackling this issue by whatever means provides hope for the future.
Make a Comment
To make a comment you must login or register.