CryoEM γ-Secretase Structures Nail APP, Notch Binding
Quick Links
Targeting the γ-secretase protease to prevent Alzheimer’s disease would require surgical precision to shut down Aβ42 production while allowing the non-pathogenic processing of amyloid precursor protein (APP) and other important substrates to continue. Researchers just took two giant steps toward that effort with the publication of not one, but two, structures of the γ-secretase complex interacting with key substrates. In the January 11 Science, Yigong Shi, Tsinghua University, Beijing, and colleagues present a high-resolution cryo-electron microscopy structure of γ-secretase enfolding a fragment of APP. This follows their December 31 report in Nature of a γ-secretase-Notch structure. The two substrates engage the enzyme in broadly similar ways, but subtle differences may open the door for substrate-specific inhibitors. The majority of AD-causing mutations in presenilin, the catalytic subunit of γ-secretase, and those in APP that could be seen in the structure, cluster at the interface between enzyme and substrate, reinforcing the idea that altered substrate binding and processing are fundamental in the genesis of AD.
- First structures of γ-secretase bound to APP reveal that transmembrane helices unwind before being cleaved.
- Small differences in binding of APP and Notch hint at potential for specific inhibitors.
- AD mutations cluster at interfaces between APP, presenilin.
Several commenters called the work a tour de force, with important implications for both familial (FAD) and late-onset AD. “The new structures provide a powerful platform for formulating specific models of normal enzyme function as well as pathological function in FAD that can be tested in biochemical experiments. Up to now, the field has been collectively operating blindfolded, generating models of substrate interaction and processing through use of chemical probes and mutagenesis. Now such experiments can be designed and interpreted with eyes wide open,” wrote Michael Wolfe, University of Kansas, Lawrence, to Alzforum.
Because it controls the spigot on Aβ42 production, γ-secretase offers a prime therapeutic target. The enzyme takes a transmembrane fragment of the amyloid precursor protein (APP) and cleaves it to produce either Aβ48 or Aβ49. From there, additional sequential bites liberate Aβ45/42/38 or Aβ46/43/40. However, the enzyme processes more than 50 substrates, including Notch, an important cell signaling receptor. Early γ-secretase inhibitors made it to Phase 3 testing, but failed when they caused worsening cognition and other side effects, including skin cancer, possibly due to inhibition of Notch cleavage or disruption of APP function (Aug 2010 news; Oct 2017 news). Since then, researchers have turned their attention to γ-secretase modulators: compounds that can selectively inhibit Aβ42 production, while allowing non-amyloidogenic processing of APP, and other substrates (for example, see Wagner et al., 2017).
Helical Home. On the left, a strand of APP occupies the transmembrane pore of PS1. Catalytic site residues are in red. A slice through the membrane (right) reveals how APP slips in on the convex side of PS1, surrounded by PS1 transmembrane helices 2, 3, 5, 6 and 7. [Courtesy of Zhou et al., Science/AAAS.]
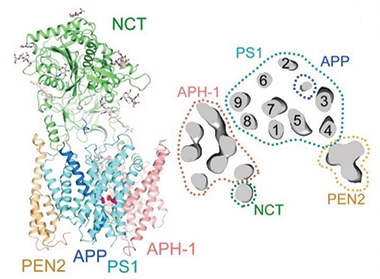
Three years ago, Shi and colleagues unveiled the first cryoEM structure of the multi-subunit γ-secretase, complete with the catalytic presenilin 1 (PS1) subunit, plus accessory proteins APH-1, PEN-2, and nicastrin (Aug 2015 news). However, enzyme-substrate complexes resisted purification. To get the flighty proteins to stick together, first authors Rui Zhou and Guanghui Yang introduced a chemical cross-link between the substrates and the extracellular loop 1 in PS1, which is thought to be involved in substrate binding. They also mutated the catalytic site to kill the protease activity, further stabilizing substrate binding. As model substrates, the researchers chose APP C83, the transmembrane fragment produced by α-secretase cleavage. Although it is not the precursor to Aβ—that would be the longer C99 product of β-secretase cleavage—C83 bound to the γ-secretase tightly and cross-linked more efficiently than C99. For Notch, they used a 100 amino acid fragment of that protein analogous to APP-C99. They were able to resolve the γ-secretase/C83 structure to an average resolution of 2.6 Å and γ-secretase/Notch-100 to 2.7 Å.

Opportune Opening. Binding to γ-secretase induces unwinding of the APP transmembrane helix, exposing two peptide bonds for cleavage. [Courtesy of Zhou et al., Science/AAAS.]
Sans substrate, γ-secretase assumes a rather loose comportment, likely existing in several conformations (Bai et al., 2015). The addition of substrate changed that, bringing order to several regions, including the highly flexible second transmembrane domain of PS.
More surprising were the contortions of APP-C83. As expected, the substrate traversed the plasma membrane in the central γ-secretase pore, surrounded by five PS1 transmembrane helices. Unexpectedly, binding caused unwinding of the final C-terminal turn of the C83 transmembrane helix, exposing the peptide bonds between APP amino acids 719/720 and 720/721 (see image above). The distal, unwound C83 residues 722-725 participated in an unusual β-sheet that contained one strand from APP and two from the cytoplasmic loop 2 of PS1. The β strand of APP also interacted with the Pro-Ala-Leu sequences at positions 433-435 of PS1. This PAL motif, previously implicated in substrate binding, helped stabilize the APP β-sheet and orient the scissile peptide bonds near the PS1 catalytic site.
The same hybrid β-sheet formed with Notch-100.
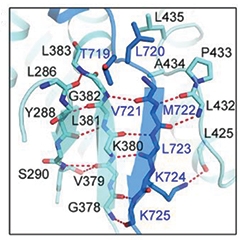
Three Strands. APP (blue) binding to γ-secretase (cyan) induces a three-strand β-sheet adjacent at the catalytic site, made up of two PS1 strands and one from APP. The PS1 P-A-L motif at residues 433-435 (upper right) helps stabilize the structure. [Courtesy of Zhou et al., Science/AAAS.]
“This structure provides a long-sought answer to the question of how the α-helix structure of the transmembrane domain is cleaved,” wrote Taisuke Tomita, University of Tokyo, to Alzforum. “We now confirm that the helix of APP/Notch is unwound in the enzyme. Dynamic conformational changes in both enzyme and substrate are critical to the intramembrane proteolysis,” he wrote.
The ability to form the hybrid β-sheet was indeed critical to enzyme activity. Deletion of residues encompassing either of the PS1 β-sheets, or disrupting the PS1 PAL sequence, destroyed PS activity toward APP-C3 or Notch-100 in vitro, the researchers showed. A key stabilizing residue for this structure, PS1 E280, is often mutated in people with early onset AD.
Yueming Li of Memorial Sloan Kettering Cancer Center in New York said that the hybrid β-sheet formation may explain why γ-secretase has so many substrates. “Any substrate with a transmembrane helix and an N-terminal flexible region could potentially be cleaved by γ-secretase,” he told Alzforum.
The structure supports a previously proposed model for how γ-secretase sequentially trims Aβ48 or 49 to shorter species (Szaruga et al., 2017). The authors speculate that once the initial cleavage occurs, and the APP C-terminal residues are released, the next turn of the helix unwinds. The unwound tail then assumes the induced β-strand structure, exposing the next cleavage site. APP may be more susceptible to sequential processing because it has more helix-destabilizing residues in its transmembrane domain to promote unwinding, compared with Notch, which has more helix-stabilizing residues, the authors speculate.
With structure in hand, the researchers considered the effect of AD-associated mutations: The majority of PS FAD mutations appeared to interfere with substrate binding or cleavage, either directly or indirectly. Many occurred in regions that contact the APP transmembrane helix or participate in forming the β-strand structure. For APP, the researchers could discern the orientation of 11 of 17 known mutations: Nine sat clustered around the cleavage site, either in the carboxyl terminal of the transmembrane helix or in the β-strand region.

Tight Pocket. Because the side chains of APP-C83 (blue) take up more space than those of Notch-100 (orange) near the active site of PS1, a small molecule may be found that specifically displaces the larger substrate. [Courtesy of Zhou et al., Science/AAAS.]
Based on the two structures, the design of substrate-specific inhibitors will be challenging, the authors wrote. APP-C83 and Notch-100 engage γ-secretase in the same location, and are stabilized by the same interactions. However, some differences emerged. In some places the APP transmembrane helix takes up less space in the PS pore than Notch, while in others places the opposite occurs. Close to the active site, bulkier APP residues might allow for APP-specific inhibitors (see image at left).
Even if medicinal chemists could turn those tiny differences into a Notch-sparing inhibitor, that may not be sufficient for an Alzheimer’s therapeutic, said Eric Siemers, formerly of Lilly and now an independent consultant. “The science is really impressive, and the authors are very good about making suggestions about how to place small molecules that may inhibit Notch cleavage or APP, and leave the other alone. However, we don’t know what caused the problems with γ-secretase inhibitors in the clinic. We don’t know whether it was [inhibition of] Notch, or another of γ-secretase’s many substrates.”
More recently, several γ-secretase modulators have been shown to reduce the production of Aβ42 selectively. Their mechanism of action is unknown, but they appear to activate the trimming activity of the γ-secretase. Recent work suggests one modulator affects the structure of the extracellular loop and first transmembrane domain of PS1 (Jul 2014 news; Cai et al., 2017). Unfortunately, the cryoEM structure doesn’t provide any additional insight. “Further structural investigations of γ-secretase with specific modulators may provide more information for design or optimization of modulators,” Yang wrote to Alzforum. Manuel Hitzenberger, Technical University of Munich, noted that stabilizing substrate binding to the enzyme complex could promote production of shorter forms of Aβ (see comment below). Yang thinks the cryoEM structure will provide a basis for studying this idea. They’re also planning to compare structures containing different presenilins and APH1 isoforms, he wrote.
Lastly, Lei Liu and Dennis Selkoe, Brigham and Women’s Hospital, Boston, caution against assuming that the binding of the longer, amyloidogenic C99 substrate will be identical to C83, as the authors hypothesize. The considerably stronger binding of C83 and the known differential processing of these substrates suggests that the precise binding interfaces may differ, they write (Jun 2017 news). In addition, they noted that the chemical crosslink introduced to stabilize the enzyme-substrate complex falls in a hotspot for PS1 FAD mutations, and it remains to be seen if that alters the structure (see comment below).—Pat McCaffrey
References
News Citations
- Lilly Halts IDENTITY Trials as Patients Worsen on Secretase Inhibitor
- Semagacestat, a Pseudo γ-Secretase Inhibitor?
- γ-Secretase Revealed in Atomic Glory
- Scientists Pinpoint γ-Secretase Modulator Binding Spot
- Substrate Says—APP Fragments Determine Where γ-Secretase Cuts
Mutation Position Table Citations
Paper Citations
- Wagner SL, Rynearson KD, Duddy SK, Zhang C, Nguyen PD, Becker A, Vo U, Masliah D, Monte L, Klee JB, Echmalian CM, Xia W, Quinti L, Johnson G, Lin JH, Kim DY, Mobley WC, Rissman RA, Tanzi RE. Pharmacological and Toxicological Properties of the Potent Oral γ-Secretase Modulator BPN-15606. J Pharmacol Exp Ther. 2017 Jul;362(1):31-44. Epub 2017 Apr 17 PubMed.
- Bai XC, Rajendra E, Yang G, Shi Y, Scheres SH. Sampling the conformational space of the catalytic subunit of human γ-secretase. Elife. 2015 Dec 1;4 PubMed.
- Szaruga M, Munteanu B, Lismont S, Veugelen S, Horré K, Mercken M, Saido TC, Ryan NS, De Vos T, Savvides SN, Gallardo R, Schymkowitz J, Rousseau F, Fox NC, Hopf C, De Strooper B, Chávez-Gutiérrez L. Alzheimer's-Causing Mutations Shift Aβ Length by Destabilizing γ-Secretase-Aβn Interactions. Cell. 2017 Jul 27;170(3):443-456.e14. PubMed. Correction.
- Cai T, Yonaga M, Tomita T. Activation of γ-Secretase Trimming Activity by Topological Changes of Transmembrane Domain 1 of Presenilin 1. J Neurosci. 2017 Dec 13;37(50):12272-12280. Epub 2017 Nov 8 PubMed.
Further Reading
Papers
- Voytyuk I, De Strooper B, Chávez-Gutiérrez L. Modulation of γ- and β-Secretases as Early Prevention Against Alzheimer's Disease. Biol Psychiatry. 2018 Feb 15;83(4):320-327. Epub 2017 Aug 10 PubMed.
Primary Papers
- Zhou R, Yang G, Guo X, Zhou Q, Lei J, Shi Y. Recognition of the amyloid precursor protein by human γ-secretase. Science. 2019 Feb 15;363(6428) Epub 2019 Jan 10 PubMed.
- Yang G, Zhou R, Zhou Q, Guo X, Yan C, Ke M, Lei J, Shi Y. Structural basis of Notch recognition by human γ-secretase. Nature. 2019 Jan;565(7738):192-197. Epub 2018 Dec 31 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Brigham and Women's Hospital
Co-Director, Brigham and Women's Hospital's Ann Romney Center for Neurologic Diseases
All readers of Zhou et al. and the companion article by Yang et al. will no doubt agree that this work from Yigong Shi’s group represents a tour de force in applying cryoEM to the structure of the γ-secretase complex with either of two key substrates cross-linked to it. The atomic resolution detailing the juxtaposition of many specific PS1 residues to those of the C83 fragment of APP or to an analogous transmembrane (TM) fragment of Notch elegantly extends certain biochemical studies of presenilin structure-function relationships reported over the last two decades. Some substrate-enzyme interactions previously postulated in biochemical analyses are now firmly established by the structural work. Two examples are the importance of the PS1 transmembrane domain (TMD) 1-2 loop in substrate recognition (Takagi-Niidome et al., 2015) and the pattern of the S1'-S2'-S3' pockets on the PS1 enzyme that respectively accept large-small-large side chains of APP to effect the tripeptide cleavages (Bolduc et al., 2016). Also now confirmed is the longstanding hypothesis that many AD-causing mutations in PS1 are aligned along the sides of the TM helices facing the TM domain of APP; the structure now depicts precisely how some of these PS1 mutations alter the interaction with APP, including with its di-aspartate active site.
A key insight from the new work that was conjectured but not mechanistically understood is the requirement for an unwinding of the substrate’s distal TM helix to expose its scissile bonds to the PS1 active site and allow hydrolysis. Zhou et al. show that C83 unwinding occurs via the formation of a 3-stranded hybrid β-sheet composed of two β-strands of PS1 (one from the distal N-terminal fragment and one from the proximal C-terminal fragment) with a new β-strand that accompanies the unwinding of one helical turn (aa 718-721) at the C-terminus of C83. This focal unwinding of the helix upon its binding to PS1 allows the epsilon cleavage to occur either at Thr719-Leu720 (yielding Aβ48) or at Leu720-Val721 (yielding Aβ49). The cross-linked structure thus explains the first (epsilon) cleavage of APP, but it cannot pinpoint the presumably analogous conformational changes underlying the subsequent tri- and tetra-peptide cleavages (“processivity”) identified biochemically by Ihara and colleagues.
A related structural insight is evidence for the binding of Met51 into a hydrophobic pocket on PS1 consistent with the large-small-large sizes of S1'-S2'-S3' pockets predicted by APP TM mutagenesis. Met51 entering the S2' pocket will lead to the 49→40 pathway, supporting more Aβ40 than Aβ42 production.
Zhou et al. hypothesize that the C99:γ-secretase structure (not yet done) may be identical to the C83:γ-secretase interactions they describe. Perhaps one should be cautious here. The authors cite a considerably stronger binding of C83 than C99 with the γ-complex, implying somewhat different binding. Moreover, the ratios of Aβ42 to Aβ40 and AICD49–99 to AICD50–99 differ according to which APP CTF substrate is being processed: C89 (β') > C99 (β) > C83 (α) (Siegel et al., 2017), suggesting that the precise E:S interfaces for these APP substrates may differ.
As the authors point out, a caveat about the interpretation of the structure of this C83-PS1 complex is the placement of the cysteine mutant into presenilin (Q112C) to achieve cross-linking. Although both covalent cross-linking and mutating one of the active site aspartates (yielding a “catalytically dead” enzyme) were necessary to stably embed C83 into γ-secretase, Q112C might not allow full representation of the dynamics of γ-secretase. Hydrophilic loop 1 (HL1) between TM1 and TM2 of PS1 (site of Q112) is involved in substrate gating (Takagi-Niidome et al., 2015) as well as in the binding sites of certain γ-secretase modulators (Cai et al., 2017). Moreover, HL-1 is a hot spot for FAD mutations (23 mutations among 34 residues), which indicates a higher vulnerability for conformational change upon residue substitution. These considerations raise a potential influence of placing the cysteine in this particular position on the described structure.
Therapeutically, the new structural details around the PS1 HL-1 loop in the E-S complex could accelerate the design of more potent and well-targeted γ-secretase modulators (GSMs), which have theoretical and practical advantages over γ-secretase inhibitors.
Finally, another complexity about APP-γ interactions arises from the recent isolation of an endogenous high MW complex of BACE1 and γ-secretase from human brain that can mediate the sequential cleavages of holoAPP to its final products, including a full array of Aβ peptides with physiological 42/40 ratios (Liu et al., 2019). An analogous high MW complex between α-secretase (ADAM10) and γ-secretase was previously described (Chen et al., 2015).
These findings predict the existence of complex interactions between BACE1, γ-secretase, and their holo-substrates that precede the formation of the C83 or C99 complexes with γ-secretase analyzed by Zhou et al.
References:
Takagi-Niidome S, Sasaki T, Osawa S, Sato T, Morishima K, Cai T, Iwatsubo T, Tomita T. Cooperative roles of hydrophilic loop 1 and the C-terminus of presenilin 1 in the substrate-gating mechanism of γ-secretase. J Neurosci. 2015 Feb 11;35(6):2646-56. PubMed.
Bolduc DM, Montagna DR, Seghers MC, Wolfe MS, Selkoe DJ. The amyloid-beta forming tripeptide cleavage mechanism of γ-secretase. Elife. 2016 Aug 31;5 PubMed.
Siegel G, Gerber H, Koch P, Bruestle O, Fraering PC, Rajendran L. The Alzheimer's Disease γ-Secretase Generates Higher 42:40 Ratios for β-Amyloid Than for p3 Peptides. Cell Rep. 2017 Jun 6;19(10):1967-1976. PubMed.
Cai T, Yonaga M, Tomita T. Activation of γ-Secretase Trimming Activity by Topological Changes of Transmembrane Domain 1 of Presenilin 1. J Neurosci. 2017 Dec 13;37(50):12272-12280. Epub 2017 Nov 8 PubMed.
Liu L, Ding L, Rovere M, Wolfe MS, Selkoe DJ. A cellular complex of BACE1 and γ-secretase sequentially generates Aβ from its full-length precursor. J Cell Biol. 2019 Feb 4;218(2):644-663. Epub 2019 Jan 9 PubMed.
Chen AC, Kim S, Shepardson N, Patel S, Hong S, Selkoe DJ. Physical and functional interaction between the α- and γ-secretases: A new model of regulated intramembrane proteolysis. J Cell Biol. 2015 Dec 21;211(6):1157-76. PubMed.
Sloan-Kettering Institute
Work in both papers represents a milestone in the study of γ-secretase and intramembrane proteases. Their findings show the similarity and differences of APP and Notch interaction with the active site of γ-secretase, which greatly advances our understanding of the specificity of γ-secretase and should help the development of effective γ-secretase based therapies.
The University of Tokyo
I’m very surprised and impressed how the authors performed experiments carefully to reveal the structure of the active γ-secretase complexed with its substrate. The key point was cross-linking of APP/Notch substrates with hydrophilic loop1 of presenilin, together with mutation at the catalytic aspartate. Using these techniques, they successfully obtained a snapshot of γ-secretase in action.
There are several surprises in these structures. First, both enzyme and substrate significantly altered their structure upon binding. In the γ-secretase, substantial changes at loop1, transmembrane domains 2/3, and TM6 were observed. In addition, the structure of the cytosolic side of the substrate was also changed to form a β-strand, which forms a hybrid β-sheet with PS1. This structure gives an answer for an age-old question regarding γ-secretase; how is the α-helix structure of the TMD cleaved? From this structure, we now confirm that the structure of APP/Notch unwinds in the enzyme. Dynamic conformational changes in both enzyme and substrate are critical to the intramembrane proteolysis.
Second, several TMs in PS1, the catalytic center of the γ-secretase, are involved in the recognition of the substrate; Loop1, TM2, TM3, TM5, TM6, TM7, and a PAL motif. These results are highly reminiscent of results of biochemical experiments by us and the others (Sato et al., 2006; Sato et al., 2008; Watanabe et al., 2010; Takagi-Niidome, 2015; Tominaga et al., 2016; Cai et al., in preparation). Moreover, they found that several FAD-linked mutations in PS1 are aligned at putative substrate binding and delivery domains, or involved in the substrate-induced conformational change. Again, global and dynamic structural alterations in PS1 are important to cleave the TM helix. FAD mutations might cause the partial loss of function by inhibition of conformational change and interaction with the substrate (Chávez-Gutiérrez et al., 2012; Szaruga et al., 2017).
Third, the binding mode of APP and Notch to the γ-secretase are essentially the same. Both substrates evoked similar conformational changes in PS1, and bound to similar regions. The authors discussed the possibility of developing selective small compound inhibitors given that the APP segment that associates with PS1 is bulkier than the Notch equivalent. However, the effect on the other substrates remains unknown. This information suggests that design of the substrate selective inhibitor based on these structures is not easy. In other words, substrate selectivity should be achieved before the incorporation of the substrate into PS1.
In fact, reduction of aggregation-prone Aβ can be achieved by γ-secretase modulators, which essentially activated the trimming activity of the γ-secretase (Takeo et al., 2014; Cai et al., 2017). We have identified that loop1 is a critical domain for the modulators, however the molecular action of these compounds is still obscure. Thus, further experiments using cryoEM would be required to understand how these modulators activate γ-secretase at the atomic level. Such information might be useful for drug development against AD.
References:
Sato C, Morohashi Y, Tomita T, Iwatsubo T. Structure of the catalytic pore of gamma-secretase probed by the accessibility of substituted cysteines. J Neurosci. 2006 Nov 15;26(46):12081-8. PubMed.
Sato C, Takagi S, Tomita T, Iwatsubo T. The C-terminal PAL motif and transmembrane domain 9 of presenilin 1 are involved in the formation of the catalytic pore of the gamma-secretase. J Neurosci. 2008 Jun 11;28(24):6264-71. PubMed.
Watanabe N, Image Image II, Takagi S, Tominaga A, Image Image I, Tomita T, Iwatsubo T. Functional analysis of the transmembrane domains of presenilin 1: participation of transmembrane domains 2 and 6 in the formation of initial substrate-binding site of gamma-secretase. J Biol Chem. 2010 Jun 25;285(26):19738-46. PubMed.
Takagi-Niidome S, Sasaki T, Osawa S, Sato T, Morishima K, Cai T, Iwatsubo T, Tomita T. Cooperative roles of hydrophilic loop 1 and the C-terminus of presenilin 1 in the substrate-gating mechanism of γ-secretase. J Neurosci. 2015 Feb 11;35(6):2646-56. PubMed.
Tominaga A, Cai T, Takagi-Niidome S, Iwatsubo T, Tomita T. Conformational Changes in Transmembrane Domain 4 of Presenilin 1 Are Associated with Altered Amyloid-β 42 Production. J Neurosci. 2016 Jan 27;36(4):1362-72. PubMed.
Chávez-Gutiérrez L, Bammens L, Benilova I, Vandersteen A, Benurwar M, Borgers M, Lismont S, Zhou L, Van Cleynenbreugel S, Esselmann H, Wiltfang J, Serneels L, Karran E, Gijsen H, Schymkowitz J, Rousseau F, Broersen K, De Strooper B. The mechanism of γ-Secretase dysfunction in familial Alzheimer disease. EMBO J. 2012 May 16;31(10):2261-74. Epub 2012 Apr 13 PubMed.
Szaruga M, Munteanu B, Lismont S, Veugelen S, Horré K, Mercken M, Saido TC, Ryan NS, De Vos T, Savvides SN, Gallardo R, Schymkowitz J, Rousseau F, Fox NC, Hopf C, De Strooper B, Chávez-Gutiérrez L. Alzheimer's-Causing Mutations Shift Aβ Length by Destabilizing γ-Secretase-Aβn Interactions. Cell. 2017 Jul 27;170(3):443-456.e14. PubMed. Correction.
Takeo K, Tanimura S, Shinoda T, Osawa S, Zahariev IK, Takegami N, Ishizuka-Katsura Y, Shinya N, Takagi-Niidome S, Tominaga A, Ohsawa N, Kimura-Someya T, Shirouzu M, Yokoshima S, Yokoyama S, Fukuyama T, Tomita T, Iwatsubo T. Allosteric regulation of γ-secretase activity by a phenylimidazole-type γ-secretase modulator. Proc Natl Acad Sci U S A. 2014 Jul 22;111(29):10544-9. Epub 2014 Jul 9 PubMed.
Cai T, Yonaga M, Tomita T. Activation of γ-Secretase Trimming Activity by Topological Changes of Transmembrane Domain 1 of Presenilin 1. J Neurosci. 2017 Dec 13;37(50):12272-12280. Epub 2017 Nov 8 PubMed.
University of Kansas
The new structures of γ-secretase bound to APP and Notch structures from the lab of Yigong Shi are together a tour de force, dramatically advancing our understanding of substrate recognition and processing by the γ-secretase complex with atomic-level detail. The conformational rearrangements of the enzyme that occur with substrate binding are substantial. In particular, several regions of the catalytic component presenilin that are disordered and unresolved in the apo structure are clearly resolved in the bound structures. Thus, these regions are apparently mobile, allowing substrate entry, whereupon they adapt by binding substrate and changing its conformation to bring it to a state suitable for peptide bond cleavage.
From the standpoint of AD, the most profound revelation is the position of presenilin FAD mutations: the large majority of these affect the interaction with APP substrate either directly or indirectly. Thus, the Shi lab has provided a structural dimension to the genetic and biochemical evidence that altered processing of APP by γ-secretase is the molecular prime mover in the pathogenesis of FAD. Such findings have clear implications for the pathogenesis of the much more common sporadic, late-onset AD, which shared the same pathology, presentation, and progression as FAD.
The new structures provide a powerful platform for formulating specific models of normal enzyme function as well as pathological function in FAD that can be tested in biochemical experiments. Up to now, the field has been collectively operating blindfolded, generating models of substrate interaction and processing through use of chemical probes and mutagenesis. Now such experiments can be designed and interpreted with eyes wide open.
Technische Universität München
The newly available cryoEM structures of the γ-secretase (GSEC)-Notch and amyloid precursor protein (APP) complexes are an important breakthrough in the ongoing investigation of GSEC substrate processing. Especially the binding mode of APP-C83 to GSEC can be expected to have an enormous impact on research regarding Alzheimer’s disease. Not only can such insight into the binding of GSEC substrates be used to determine the mechanism behind many known FAD-causing mutations, it also finally allows for rigorous structure-based approaches in drug discovery for GSEC modulation.
Our independently predicted (Hitzenberger and Zacharias, 2018) geometry is in good agreement with the newly released structures of Shi et al., validating the application of theoretical methods to GSEC-substrate interactions. In addition to the similarities with our findings, Shi et al. furthermore predict the formation of a highly stabilizing β-sheet, involving the C-terminus of the substrates. This new structural feature is highly interesting as it allows the assessment of the role of many mutations that might be involved in the determination of the cleaving pathway: It is widely believed that APP-C99 can be cleaved in two different ways: Either C99 → Aβ49 → Aβ46 → Aβ43 → Aβ40, or C99 → Aβ48 → Aβ45 → Aβ42. The latter route leads to the release of longer (i.e., more aggregation-prone) amyloids. With the newly available high-resolution structures, the role of amino acids at or close to the scissile bond of the substrate can be targeted for in-depth investigations.
It has already been shown that the stability of the enzyme-substrate complex influences the length of the Aβ product (Szaruga et al., 2017). It is therefore very likely that finding modulators that can increase the lifetime of the Aβn-GSEC complex might be a viable route to prevent the production of very aggregation-prone (i.e., long) amyloids. The novel C83-GSEC complex structure presents an invaluable starting point for any such study. Having both the Notch- and C83-GSEC complex structures available might also allow for the creation of target-specific modulators that mainly affect the processing of the desired substrate.
Since GSEC, however, processes its substrate in a succession of several cleaving steps, it would be very helpful to also know the structural properties of the cleaving intermediates (Aβ49, Aβ46, and Aβ43). This would allow the identification of ligand binding sites for modulators that aid the stabilization of the presumably less stable Aβ43(42)-GSEC complex.
Structural properties of transition states and processing intermediates are of course very challenging to investigate by experimental means and this is where theoretical approaches can be of great assistance. Since we performed computer simulations we were able to investigate the processing mechanism of APP cleavage, including all intermediate states. We therefore predict the formation of a stable Aβn-GSEC binding region formed by the substrate’s N-terminus and the nicastrin-presenilin interface, close to the C-terminus of transmembrane domain 3. Since this region stays topologically unaltered in all our simulations of the APP processing intermediates as well, it is very likely to present an interesting target for novel GSEC modulators. It is to be expected that these newly released structures in tandem with computational, mutational, and drug screening approaches, will spawn a new wave of promising small molecule ligands, targeted at modulating GSEC to produce smaller, less aggregation-prone amyloids.
References:
Hitzenberger M, Zacharias M. Structural Modeling of γ-Secretase Aβ n Complex Formation and Substrate Processing. ACS Chem Neurosci. 2019 Mar 20;10(3):1826-1840. Epub 2019 Jan 30 PubMed.
Szaruga M, Munteanu B, Lismont S, Veugelen S, Horré K, Mercken M, Saido TC, Ryan NS, De Vos T, Savvides SN, Gallardo R, Schymkowitz J, Rousseau F, Fox NC, Hopf C, De Strooper B, Chávez-Gutiérrez L. Alzheimer's-Causing Mutations Shift Aβ Length by Destabilizing γ-Secretase-Aβn Interactions. Cell. 2017 Jul 27;170(3):443-456.e14. PubMed. Correction.
K.U.Leuven and V.I.B.
I would like to start by congratulating Shi and colleagues for these recent achievements! These are exciting times for the γ-secretase field.
The recent γ-secretase-substrate structures confirm what was anticipated by the previous co-structure of the γ-secretase with a co-purifying peptide (PDB 5FN3): the substrate binding site is largely immersed in the PSEN1-NTF fragment (TM2, TM3, TM5) and extends towards the cytosolic side of the PSEN-1 NFT/CFT interface, where the two catalytic aspartates align for catalysis.
The novel structures point to conformational changes in both substrate and protease upon engaging in the Enzyme-Substrate (E-S) complex, add unprecedented atomic detail to the E-S interface, reveal an unanticipated intimate contact between the ectodomain of the substrate and the protease (Nicastrin) and uncover the critical pockets (S'1-S'3) that have been shown to modulate cleavage specificity (Bolduc et al., 2016).
Interestingly, the global conformation of the γ-secretase complex is rather similar between the two reported E-S structures, but distinct subsets of PSEN1 amino acids establish contacts with the Notch and APP substrates. The authors suggest that differential substrate recognition by γ-secretase relies on the distinct E-S interfaces and that the observed differences might be exploited for the development of substrate-specific γ-secretase inhibitors. These proposals give high functional relevance to the primary structure of the substrates (amino acid sequence), an idea that is in contrast with the large number of substrates processed by γ-secretases, presenting low sequence conservation. The development of substrate-specific inhibitors—although appealing—represents a challenging task, since the “APP-specific” inhibitors should spare the processing of many other substrates, for which no structural data is available, and even if successful, it may be a risky approach, as it will result in the accumulation of APP C-terminal fragments in the membrane and potentially lead to aberrant cell signaling and/or altered membrane homeostasis.
I am certain that the novel structural templates will facilitate functional analyses addressing the molecular mechanisms of these fascinating proteases. Interestingly, the recent structures depict substrate unwinding near the site of cleavage, indicating that this step indeed precedes γ-secretase proteolysis of helical transmembrane domains. Furthermore, they reveal a network of E-S interactions that could assist during the initial (endopeptidase) cleavage. What happens next in the sequential proteolytic process—leading to the release of N-terminal peptides of high pathophysiological relevance in case of the APP substrate—remains speculative.
In this regard, however, a previous structure of the γ-secretase with a co-purifying peptide (PDB:5FN3) provides interesting hints that guide our thoughts about how the successive processing of APP and the generation of into Aβ peptides of various length could occur. As we proposed, this structure may depict the interaction between γ-secretase and the fragment generated after the initial proteolytic cleavage of the transmembrane domain of the substrate (i.e., a “de novo”-generated long Aβ from APP), just before it engages into the next catalytic turnover (Szaruga et al., 2017). The putative substrate occupies the (now proven) substrate binding site, but in contrast to APP, only the most N-terminal part of its transmembrane domain remains in a helical structure, while its C-terminal part is unstructured and extends along the substrate binding channel to reach the catalytic residues. This implies that the first endopeptidase-mediated backbone break exerts a strong destabilizing effect on the helical structure of the generated transmembrane fragment (long Aβ from APP). This model suggests that the type of interactions established in the γ-secretase-APP vs. γ-secretase-Aβ complex are fundamentally different.
(A) PSEN1 – APP (E-S) complex. (B) Transmembrane helix unwinding occurs in order to fill the enzyme pockets during catalysis. (C) The endopeptidase-mediated backbone break destabilizes the most C-terminal part of the helical structure of the ‘de novo’ generated substrate. (D) Each sequential cleavage further unwinds the N-terminal helix (E-S anchor). This stretches the substrate, providing the length to the substrate to reach the active site. Shortening of the "helical anchor" progressively destabilizes γ-secretase-Aβ interactions, shifting the equilibrium toward dissociation (Aβ release). (A–D) PSEN1 structure (lateral view, brown) (PDB: 5FN2, [Bai et al., 2015]) with the structure of APPC99 (purple) (PDB: 2LP1, [Barrett et al., 2012]) manually docked in the putative substrate binding site.
In addition, it supports a model in which further unwinding of the N-terminal helix of the substrate must occur with each γ-secretase cut in order to provide the length of the substrate to fill the S1'–S3' enzyme pockets (Bolduc et al., 2016) during the stepwise catalysis. This “unwinding model” originally proposed in Szaruga et al., 2017 (see image), suggests also that the interactions established between the N-terminal helical structure of the substrate and the protease “anchor” the E-S complex and thereby define the length of the N-terminal product (Aβ from APP).
Finally, when it comes to the crucial role of γ-secretase in Alzheimer’s pathogenesis, I would like to highlight the fact that AD-causing mutations delineating the γ-secretase-substrate interface give structural support to our recent findings showing that AD-causing PSEN1 mutations destabilize the γ-secretase-APP/Aβ interactions and consequently enhance the release of longer Aβ peptides.
It is quite exciting to see that the different structures can come together and help us to (de)construct a complex proteolytic process!
References:
Bolduc DM, Montagna DR, Gu Y, Selkoe DJ, Wolfe MS. Nicastrin functions to sterically hinder γ-secretase-substrate interactions driven by substrate transmembrane domain. Proc Natl Acad Sci U S A. 2016 Feb 2;113(5):E509-18. Epub 2015 Dec 22 PubMed.
Szaruga M, Munteanu B, Lismont S, Veugelen S, Horré K, Mercken M, Saido TC, Ryan NS, De Vos T, Savvides SN, Gallardo R, Schymkowitz J, Rousseau F, Fox NC, Hopf C, De Strooper B, Chávez-Gutiérrez L. Alzheimer's-Causing Mutations Shift Aβ Length by Destabilizing γ-Secretase-Aβn Interactions. Cell. 2017 Jul 27;170(3):443-456.e14. PubMed. Correction.
Ludwig-Maximilians-University Munich and German Center for Neurodegenerative Diseases, Munich
The two new papers by the Shi lab are very important for the field; they clearly show that the substrate unfolds at the initial cleavage sites when it is in complex with the protease, a prerequisite required for cleavage of the α-helical substrate transmembrane domain. Both papers show similar structural features for the two major and best-studied γ-secretase substrates APP and Notch, and provide fascinating structural insights into how γ-secretase interacts with substrates. Thus, for the first time we see that upon interaction with the substrate and unfolding at the ε-sites, a β-strand is formed at the substrates’ C-terminal transmembrane domain end/membrane-anchor region, which is stabilized by two β-strands induced in the presenilin N- and C-terminal fragments (NTF, CTF) and by the PAL motif.
Both enzyme–substrate complex structures also show that a large part of the substrate transmembrane domains is surrounded by the cavity formed by presenilin transmembrane domains 2, 3 and 5. This nicely confirms an earlier study from our lab, in which we showed by introducing a photocrosslinkable amino acid into C99 at each position over two-thirds of the molecule, that the major γ-secretase substrate-binding site is the NTF (Fukumori and Steiner, 2016). It is also nice to see that the interactions of several substrate residues which we identified previously for APP C99 by this photoaffinity-mapping approach with the NTF (e.g. V44) are also seen in the conformational snapshot of the γ-secretase–APP C83 substrate complex. In addition, certain residues that are part of the β-strand of the substrate were found in our study to contact the CTF and, to a lesser extent, the NTF (e.g., L52), which is also of interest in light of the new structural data.
In our previous study, we further showed that presenilin FAD mutations can alter the interactions of the C99 cleavage domain with presenilin (Fukumori and Steiner, 2016). In follow-up structural studies, it will thus, for example, be interesting to see what the APP structure will look like in the presence of the clinical mutations. Clearly, many open questions that lie ahead will benefit now from the possibility that structural biologists can obtain high-resolution structures of γ-secretase in complex with substrate.
References:
Fukumori A, Steiner H. Substrate recruitment of γ-secretase and mechanism of clinical presenilin mutations revealed by photoaffinity mapping. EMBO J. 2016 Aug 1;35(15):1628-43. Epub 2016 May 23 PubMed.
Scripps Research
The molecular basis for differential cut sites is an interesting one.
Make a Comment
To make a comment you must login or register.