Cutting (or Slippery?) Edge: Lipids in Neurodegeneration Science
Quick Links
Readers, ignore lipids at your peril. These oily molecules make up 60 percent of the human brain. Long considered mere wrapping for more interesting cellular contents, these fats hold secrets worth exploring. At the 2nd Symposium on Lipids in Brain Diseases, held September 13-15 in Leiden, The Netherlands, about 100 experts on lipids mingled with nonexperts to share data and ideas in discussions of how lipids both grease and prevent neurological disease.
- Second Lipids in Brain Disease Symposium held in Leiden.
- At its forefront: what role fatty acids?
- Their dysregulation may hasten α-synuclein pathology.
- The same lipids might protect against ALS/FTD.
“It’s the new frontier,” said Adrian Isaacs, University College London. Saranna Fanning, Brigham and Women’s Hospital, Boston, said lipids have potential as biomarkers, drug targets, and therapeutic strategies. “We are at the point where the study of brain lipids is worth multiple meetings a year,” she told Alzforum.
Herman Boerhaave would have approved. Renowned for his hands-on teaching style, and sometimes called the "Dutch Hippocrates", this physician, chemist, botanist, and humanist attracted scholars from all over the world to his town in southern Holland starting in the late 17th century. Rijksmuseum Boerhaave is built around his theatrum anatomicum (image below), which stands today as it did almost 400 years ago. It was the venue for this lipid conference.

Theatrum Anatomicum. The theater stands today, just as it was when Herman Boerhaave and other scholars probed the workings of the human body in its center circle. [Courtesy of Tom Fagan.]
The organizers were Birol Cabukusta and Martin Giera from Leiden University Medical Center; Jerome Hendriks, Hasselt University, Hasselt, Belgium; Gijs Kooij, Amsterdam University Medical Center; and Rik van der Kant, Vrije University, Amsterdam.
Featuring nine keynote and nine shorter talks, the meeting was well-suited for this hot topic. Attendees learned about how different lipids operate in proteinopathies, how cells regulate myelin in health and disease, and how diet affects lipid metabolism. They also considered how the fine balance between lipids causing and resolving inflammation might open new windows for therapy.
In this four-part series, Alzforum summarizes highlights from the meeting, which will reconvene in 2025. The stories focus on imbalances in long-chain fatty acid metabolism being both a cause and a consequence of proteinopathy, on the role of microglia in maintaining the cholesterol-laden myelin sheaths, on sterols driving amyloid and tau pathology, and on how mysterious specialized lipids can help resolve inflammation.
Lipids and Proteinopathies
Scientists have known for decades that neurodegeneration and problems with lipids can go hand in hand. In lipid storage disorders, such as Nieman Pick’s disease, cholesterol and sphingolipids clog lysosomes, preventing cells from degrading and recycling unwanted protein and leading to the death of neurons For a good synopsis, see Pfrieger 2023.
By now, lipids have come under intense scrutiny in Alzheimer’s, Parkinson’s, and other neurodegenerative disorders. Genome-wide association studies have linked lipid metabolism genes to AD. Lipid droplets accumulate in microglia in the AD brain and mouse models (Aug 2019 news; Claes et al., 2021; Sep 2023 news).
In Parkinson’s, Fanning, when working in Dennis Selkoe’s lab at Brigham and Women’s, had previously linked changes in the lipid bilayer of cellular membranes to aggregation of α-synuclein, the principal component of Lewy bodies found in PD and other synucleinopathies, including dementia with Lewy bodies.
Specifically, Fanning reported an exacerbation of α-synuclein pathology, thanks to the enzyme stearoyl CoA desaturase. SCD introduces a double bond into stearic acid, an 18-carbon saturated fatty acid, to create oleic acid, a monounsaturated 18C fatty acid. This cis double bond, between carbon 9 and 10, forces a 30° kink in the fatty acid chain. This loosens the packing of the fatty acid and its derivatives, such as phosphatidylcholine, in lipid membranes. As a result, monomers of α-synuclein can linger in the membrane a little longer than usual, giving them more opportunity to aggregate, Fanning found. Blocking SCD attenuated α-synuclein aggregation and toxicity in yeast, roundworms, and human neurons (Dec 2018 news).

Tri This. Three fatty acid chains attach to the 3-carbon glycerol backbone to form triglycerides. In this case the fatty acids are palmitic (top), oleic (middle), and α-linolenic. In mammals, fatty acids are stored and transported as triglycerides.
If oleic acid spells trouble, then would it be wise to rid cells of excess, including any oleic acid stored in triglycerides? Not exactly. In Leiden, Fanning reported that blocking metabolism of these fatty acid troikas (image at right) can also alleviate synuclein toxicity. When she blocked lipase E (LIPE) in neuroblastoma cells expressing mutant α-synuclein, fewer aggregates formed and less of the protein was phosphorylated on serine 129, a marker of α-synuclein toxicity.
LIPE hydrolyses neutral lipids, specifically the di- and triglycerides that accumulate in lipid droplets. In so doing, this enzyme releases free fatty acids for further metabolism. Knocking down its expression with a short hairpin RNA also protected the cells from α-synuclein toxicity. Likewise, knocking down the lipase protected dopaminergic neurons from degenerating in a C. elegans model of α-synuclein toxicity.

As You LIPE It. Cells expressing mutant -synuclein (green) accumulate aggregates of the protein (left), but not if a lipase is blocked (right). [Courtesy of Fanning et al., 2022.]
How does silencing LIPE help? The free fatty acids it liberates from lipid droplets and triglycerides include oleic acid—that same one made by the enzyme SCD (image at right). Fanning thinks cells cannot metabolize this fatty acid fast enough once triglycerides have been hydrolyzed, leaving it free to slither into cell membranes where it promotes aggregation of α-synuclein. In short, it might be better to leave oleic acid sequestered away in the lipid droplets, where it does less harm.
What might be the better approach—stopping the synthesis of oleic acid, or its release from triglycerides? Perhaps the answer is both. “Ideally, you would prevent the synthesis,” Fanning said, “but by the time a patient enters the physician's office, the synthesis pathway may have been cranking out oleic acid for over a decade, and lipases may be releasing any stored in lipid droplets. So if we reduce LIPE activity at that stage, it may be helpful.”
Indeed, in Leiden Fanning reported that, at least in neuroblastoma cells, inhibiting both the making and the breaking enzymes, i.e. SCD and LIPE, beat blocking either alone at reducing levels of oleic acid, p-Syn129, and α-synuclein aggregates.
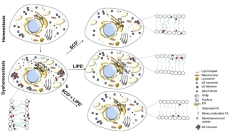
Lipid Balance. Healthy cells (top left) have few di- and triglycerides, lipid droplets, and α-synuclein aggregates. Not so when mutant α-synuclein is around (bottom left). Blocking SCD to limit monosaturated fatty acids (top right), blocking LIPE to prevent degradation of di- and triglycerides (center right), or both (bottom right) might hold off synuclein toxicity. [Courtesy of Fanning et al., 2022.]
Blocking LIPE could have downsides. For one, lipid droplets might accumulate. “When manipulating any lipid in cells, you want to be sure to return membranes to the fatty acid composition associated with health,” said Fanning. One way to do this might be to counter monounsaturated fatty acids, such as oleic acid, with saturated ones.
Ulf Dettmer at Brigham and Women’s Hospital, who collaborates with Fanning, has done just that. He loaded neuroblastoma cells with 14-carbon myristic acid, 16-carbon palmitic acid, and 18-carbon stearic acid. All three tempered phosphorylation of α-synuclein at serine 129, while myristic acid increased the multimeric-to-monomeric ratio of α-synuclein and decreased aggregates (Imberdis et al., 2019). α-Synuclein tetramers are believed to be more stable and less likely to misfold into fibrils (Aug 2011 news). Together, Dettmer and Fanning are currently exploring how myristic and other fatty acids affect α-synuclein toxicity.
Others in Leiden called the work impressive. “This is a good example of how scientists can delve into lipidomics and come up with potential drug targets,” van der Kant said. “When we go to clinical trial meetings we never hear people talking about lipids, but this is an example of how they may help tackle pathology,” he said.
Femke Feringa, also from Vrije University, noted that lipid research in the past has been held back by the difficulties of understanding how the fluidity of membranes affects biological processes. “Finding the best systems to study this relationship is a big challenge” she said. “Now that we have iPSC models, we can hopefully find better experimental approaches.” For more on that, see below.
What about other proteinopathies? In Leiden, UCL’s Isaacs described quite a different instance of lipid dysregulation, though still of the fatty acid kind. Isaacs studies models of amyotrophic lateral sclerosis/frontotemporal dementia.
Scientists in his lab developed a fly model of ALS/FTD based on expression of the dipeptide repeats that are translated from the CCGGG hexanucleotide expansion in the C9ORF72 gene (Mizielinska et al., 2014).
Since then, Ashling Giblin in the lab has found that, of the top 10 diminished processes in these flies, three had to do with lipid synthesis. Downregulated genes included acetyl-coenzyme A synthetase, which makes the metabolite acetyl CoA, and acetyl CoA carboxylase, which turns acetyl CoA into malonyl CoA. Malonyl CoA is the rate-limiting substrate in the formation of longer-chain fatty acids.
Expression of FASN1 and FASN2 genes, which encode enzymes that make those long-chain fatty acids from malonyl CoA, were also downregulated. So was SCD1, the desaturase that turns stearic acid into oleic acid. C9ORF72 flies made half as much FASN1 and SCD1 RNA as did controls. This jibes with prior transcriptomic analysis of postmortem ALS spinal cord. Among 154 people with the disease, FASN1 expression was almost halved, and SCD1 expression was down about 10 percent, compared to control tissue (Humphrey et al., 2023).
What about human cells? Alex Cammack in Isaacs' lab found that there is a dearth of highly unsaturated phospholipids in neurons generated from iPSC cells carrying a C9ORF72 expansion. Expressing a 92-repeat hexanucleotide—the length that gives a person disease—in normal neurons tanked the same phospholipids, whereas adding antisense nucleotides to silence the expansions restored lipid balance.
Conceptually, could restoring healthy lipid profiles protect against the C9ORF72 repeat? In the fly model, overexpressing FASN1 or SCD1 extended lifespan, while in the human C9ORF72 neurons, overexpressing enzymes that generate polyunsaturated fatty acids increased cell viability “We see a very specific change in phospholipids that contain highly saturated fatty acids; if we can reverse that therapeutically, it may have great potential,” Isaacs told the audience.
The literature hints that he might be right. In a prospective study of a million people in the U.S., those who ate more ω3 polyunsaturated fatty acids, such as the 18-carbon, were less likely to develop ALS in the next two decades (Fitzgerald et al., 2014). The ω3 PUFAs are those whose first double bond occurs at the third carbon from the methyl end of the chain. Two case-control studies had also linked high PUFA intake to reduced risk for the disease (Veldink et al., 2007; Okamoto et al., 2007). More direct evidence has come from analysis of plasma. In a Phase 3 clinical trial of dexpramipexole in ALS, people who had more of the ω3 eicosapentaenoic acid and the ω6 linoleic acid at the beginning of the trial were likelier to still live 12 months later (Bjornevik et al., 2023).
Manipulating brain lipids through the diet may be a tall order, since many of these lipids bounce off the blood-brain barrier. Indeed, Isaacs found that while feeding PUFAs to the C9ORF72 flies made them live longer, this was less effective than expressing the enzymes that make unsaturated fatty acids.
Attendees in Leiden found Isaac’s data fascinating. Tiago Gil Oliveira, Universidade do Minho, Portugal, asked if the lipid imbalance affects the fluidity of cell membranes. Isaacs said this question was top of his mind these days. Others wondered how the dipeptide repeats made from the expanded C9ORF72 RNA change lipid dynamics. Isaacs said this needs to be figured out.—Tom Fagan
References
News Citations
- Newly Identified Microglia Contain Lipid Droplets, Harm Brain
- Lipid-Laden, Sluggish Microglia? Blame Aβ.
- Does Oleic Acid Hold the Key to α-Synuclein Toxicity?
- An α-Synuclein Twist—Native Protein a Helical Tetramer
Therapeutics Citations
Paper Citations
- Pfrieger FW. The Niemann-Pick type diseases - A synopsis of inborn errors in sphingolipid and cholesterol metabolism. Prog Lipid Res. 2023 Apr;90:101225. Epub 2023 Mar 31 PubMed.
- Claes C, Danhash EP, Hasselmann J, Chadarevian JP, Shabestari SK, England WE, Lim TE, Hidalgo JL, Spitale RC, Davtyan H, Blurton-Jones M. Plaque-associated human microglia accumulate lipid droplets in a chimeric model of Alzheimer's disease. Mol Neurodegener. 2021 Jul 23;16(1):50. PubMed.
- Fanning S, Cirka H, Thies JL, Jeong J, Niemi SM, Yoon J, Ho GP, Pacheco JA, Dettmer U, Liu L, Clish CB, Hodgetts KJ, Hutchinson JN, Muratore CR, Caldwell GA, Caldwell KA, Selkoe D. Lipase regulation of cellular fatty acid homeostasis as a Parkinson's disease therapeutic strategy. NPJ Parkinsons Dis. 2022 Jun 9;8(1):74. PubMed.
- Imberdis T, Negri J, Ramalingam N, Terry-Kantor E, Ho GP, Fanning S, Stirtz G, Kim TE, Levy OA, Young-Pearse TL, Selkoe D, Dettmer U. Cell models of lipid-rich α-synuclein aggregation validate known modifiers of α-synuclein biology and identify stearoyl-CoA desaturase. Proc Natl Acad Sci U S A. 2019 Oct 8;116(41):20760-20769. Epub 2019 Sep 23 PubMed.
- Mizielinska S, Grönke S, Niccoli T, Ridler CE, Clayton EL, Devoy A, Moens T, Norona FE, Woollacott IO, Pietrzyk J, Cleverley K, Nicoll AJ, Pickering-Brown S, Dols J, Cabecinha M, Hendrich O, Fratta P, Fisher EM, Partridge L, Isaacs AM. C9orf72 repeat expansions cause neurodegeneration in Drosophila through arginine-rich proteins. Science. 2014 Sep 5;345(6201):1192-1194. Epub 2014 Aug 7 PubMed.
- Humphrey J, Venkatesh S, Hasan R, Herb JT, de Paiva Lopes K, Küçükali F, Byrska-Bishop M, Evani US, Narzisi G, Fagegaltier D, NYGC ALS Consortium, Sleegers K, Phatnani H, Knowles DA, Fratta P, Raj T. Integrative transcriptomic analysis of the amyotrophic lateral sclerosis spinal cord implicates glial activation and suggests new risk genes. Nat Neurosci. 2023 Jan;26(1):150-162. Epub 2022 Dec 8 PubMed.
- Fitzgerald KC, O'Reilly ÉJ, Falcone GJ, McCullough ML, Park Y, Kolonel LN, Ascherio A. Dietary ω-3 polyunsaturated fatty acid intake and risk for amyotrophic lateral sclerosis. JAMA Neurol. 2014 Sep;71(9):1102-10. PubMed.
- Veldink JH, Kalmijn S, Groeneveld GJ, Wunderink W, Koster A, de Vries JH, van der Luyt J, Wokke JH, Van den Berg LH. Intake of polyunsaturated fatty acids and vitamin E reduces the risk of developing amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry. 2007 Apr;78(4):367-71. Epub 2006 Apr 28 PubMed.
- Okamoto K, Kihira T, Kondo T, Kobashi G, Washio M, Sasaki S, Yokoyama T, Miyake Y, Sakamoto N, Inaba Y, Nagai M. Nutritional status and risk of amyotrophic lateral sclerosis in Japan. Amyotroph Lateral Scler. 2007 Oct;8(5):300-4. PubMed.
- Bjornevik K, Cortese M, Furtado JD, Paganoni S, Schwarzschild MA, Cudkowicz ME, Ascherio A. Association of Polyunsaturated Fatty Acids and Clinical Progression in Patients With ALS: Post Hoc Analysis of the EMPOWER Trial. Neurology. 2023 Aug 15;101(7):e690-e698. Epub 2023 Jun 21 PubMed.
Further Reading
No Available Further Reading
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.