TDP-43 Structure Reveals Two-Faced Amino End
Quick Links
The amino end of TDP-43 has evaded structural characterization because of its penchant for aggregation. Now it has finally revealed its shape. Two shapes, in fact: The domain flips between a well-folded nucleic acid-binding conformation and a loose, aggregation-prone jumble, according to a paper in the Proceedings of the National Academy of Sciences this week. Unexpectedly, the folded conformation looks much like ubiquitin, but this version binds nucleic acids instead of proteins. The authors from the National University of Singapore propose that the folded configuration could work in the nucleus, where TDP-43 regulates gene expression, while the floppy disarray likely promotes cytoplasmic aggregation. Small molecules that lock the domain in the more steadfast form might be therapeutic for TDP-43 proteinopathies, which include amyotrophic lateral sclerosis and frontotemporal dementia, suggested senior author Jianxing Song in an email to Alzforum.
In addition to the amino-terminal domain, TDP-43 contains two RNA-binding motifs and a glycine-rich carboxyl terminus. The majority of disease-linked mutations cluster in the carboxyl end, so scientists have focused their attention on that section, noted Magdalini Polymenidou of the University of Zürich, who was not involved in the study. However, last year researchers at the Mayo Clinic in Jacksonville, Florida, reported that the protein’s amino tip mediates both TDP-43’s function in RNA splicing, and its aggregation (Zhang et al., 2013). Song’s dual structures reveal how the amino end might be involved in these processes. “It really shows that we have overlooked the amino terminus,” Polymenidou said. “We need to study the entire protein.”

Two of a kind.
The structure of the TDP-43 amino end (purple) neatly aligns with that of ubiquitin (yellow). [Image courtesy of Qin et al., PNAS.]
TDP-43, like many proteins involved in neurodegeneration, confounds structural biologists with its aggregation, Song wrote. In TDP-43, the amino terminus has the highest propensity to cluster, he added. Song and study first author Haina Qin came up with a trick to decipher the structures of these typically insoluble proteins by simply dissolving them in purified, salt-free water (Li et al., 2006). Along with co-first authors Liang-Zhong Lim and Yuanyuan Wei, they have already applied this technique to crack the structures of the ALS-related proteins SOD1 and VAPB (Lim et al., 2014; Qin et al, 2013). With the TDP-43 amino domain in pure water, the authors were able to use both nuclear magnetic resonance (NMR) spectroscopy and circular dichroism to solve its structures. They observed the disordered form in equilibrium with the tightly folded one. The more concentrated the protein, the more likely it was to adopt the floppy, aggregation-prone shape.
The stiffer version, surprisingly, folds like ubiquitin (see image above). TDP-43’s amino acid sequence provided no hints of similarity to ubiquitin; it does not even contain the lysines used by ubiquitin to connect with target proteins, noted Philipp Kahle of the University of Tübingen, Germany, who was not part of the study group. Kahle said he was initially skeptical that TDP-43 would form an ubiquitin-like fold, but found the overlay between the TDP-43 and ubiquitin shapes compelling. Other experts contacted by Alzforum also found the structural work to be sound. “The NMR data are convincing,” wrote Elizabeth Meiering of the University of Waterloo, Canada. “I think it is believable,” agreed Gregory Petsko of Weill Cornell Medical College in New York City.
Without those lysines, what does this ubiquitin copycat bind to? Since TDP-43 contains two known RNA-interacting segments, and the amino terminus was previously reported to bind DNA (Chang et al., 2012), Song and colleagues incubated the amino domain with nucleic acids to look for interactions. TDP-43 is known to bind RNAs with alternating uridines and guanines, so they tried those UGUGUG constructs as well as the corresponding single-stranded DNA with thymines and guanines. The amino terminus, when folded, bound both nucleic acids. RNA-binding proteins typically exhibit affinity for DNA as well, Polymenidou noted. Kahle suggested that in the cell, the TDP-43 amino domain would more likely bind RNA than DNA. Thousands of RNAs are known to interact with TDP-43, while only a couple of DNA sequences do so (see Nov 2010 conference story; Mar 2011 news story; Ou et al., 1995; and Lalmansingh et al., 2011).
“To the best of our knowledge, the TDP-43 amino terminus is the first ubiquitin-like fold which can directly bind nucleic acids,” Song wrote. Other proteins might contain similarly structured domains, but they would not be obvious by sequence gazing, he pointed out. Perhaps, Kahle suggested, Song’s pure-water technique will reveal other structural siblings of ubiquitin.
“I find it remarkable that you have a peptide that yields two conformers; one is wonderfully folded and the other is not,” Kahle said. Typically, proteins with alternate conformations toggle between two tightly folded states, though at least one other, a kind of sarcoma homology 3 domain, also has structured and unstructured states (Zhang et al., 1994). In the case of TDP-43, the first 25 residues of the protein are more disordered than those in ubiquitin, Song and colleagues report, which may explain its propensity to hang loose.
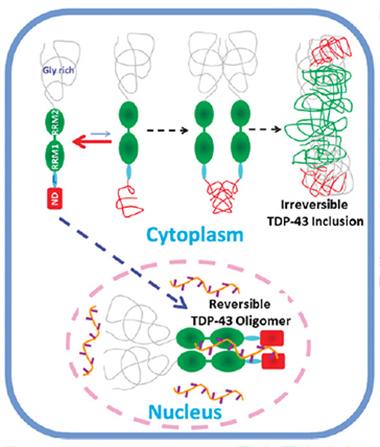
One TDP-43—Two Pathways.
Jianxing Song proposes that TDP-43 exists in equilibrium between tightly structured and loose forms. The folded version forms functional oligomers in the nucleus, where it regulates gene expression. The unfolded proteins coalesce to make inclusions in the cytoplasm. [Image courtesy of Qin et al., PNAS.]
Why would the protein contain a segment that makes it prone to unfold and aggregate? “I think it is a compromise of nature,” Kahle suggested. He speculated that the unusually loose folds of TDP-43’s amino end might allow it to be somewhat nonspecific in its binding, so it could find all those different RNA targets. However, he cautioned, this in vitro study cannot confirm that the same folded-to-unfolded switch occurs in cells.
If the two conformations do arise in cells, Song and colleagues suggest a model in which the tightly folded version attends to RNA in the nucleus, while the flexible one aggregates in the cytoplasm. The high concentration of TDP-43 in stress granules might promote the switch to the latter conformer and initiate aggregation, Polymenidou posited.
The results suggest a possible route to treat TDP-43 proteinopathies by preventing unfolding and aggregation. “We are now focused on the discovery and design of molecules to stabilize the folded form of the TDP-43 amino terminus, in an attempt to develop drug leads,” Song wrote. Polymenidou agreed that this therapeutic strategy makes sense.—Amber Dance
References
News Citations
- San Diego: TDP-43 Targets Loom Large—But Where’s the Bull’s Eye?
- CLIPs of TDP-43 Provide a Glimpse Into Pathology
Paper Citations
- Zhang YJ, Caulfield T, Xu YF, Gendron TF, Hubbard J, Stetler C, Sasaguri H, Whitelaw EC, Cai S, Lee WC, Petrucelli L. The dual functions of the extreme N-terminus of TDP-43 in regulating its biological activity and inclusion formation. Hum Mol Genet. 2013 Aug 1;22(15):3112-22. PubMed.
- Li M, Liu J, Ran X, Fang M, Shi J, Qin H, Goh JM, Song J. Resurrecting abandoned proteins with pure water: CD and NMR studies of protein fragments solubilized in salt-free water. Biophys J. 2006 Dec 1;91(11):4201-9. Epub 2006 Sep 15 PubMed.
- Lim L, Lee X, Song J. Mechanism for transforming cytosolic SOD1 into integral membrane proteins of organelles by ALS-causing mutations. Biochim Biophys Acta. 2015 Jan;1848(1 Pt A):1-7. Epub 2014 Oct 12 PubMed.
- Qin H, Lim L, Wei Y, Gupta G, Song J. Resolving the paradox for protein aggregation diseases: a common mechanism for aggregated proteins to initially attack membranes without needing aggregates. F1000Res. 2013;2:221. Epub 2013 Oct 21 PubMed.
- Chang CK, Wu TH, Wu CY, Chiang MH, Toh EK, Hsu YC, Lin KF, Liao YH, Huang TH, Huang JJ. The N-terminus of TDP-43 promotes its oligomerization and enhances DNA binding affinity. Biochem Biophys Res Commun. 2012 Aug 24;425(2):219-24. Epub 2012 Jul 23 PubMed.
- Ou SH, Wu F, Harrich D, García-Martínez LF, Gaynor RB. Cloning and characterization of a novel cellular protein, TDP-43, that binds to human immunodeficiency virus type 1 TAR DNA sequence motifs. J Virol. 1995 Jun;69(6):3584-96. PubMed.
- Lalmansingh AS, Urekar CJ, Reddi PP. TDP-43 is a transcriptional repressor: the testis-specific mouse acrv1 gene is a TDP-43 target in vivo. J Biol Chem. 2011 Apr 1;286(13):10970-82. PubMed.
- Zhang O, Kay LE, Olivier JP, Forman-Kay JD. Backbone 1H and 15N resonance assignments of the N-terminal SH3 domain of drk in folded and unfolded states using enhanced-sensitivity pulsed field gradient NMR techniques. J Biomol NMR. 1994 Nov;4(6):845-58. PubMed.
External Citations
Further Reading
Papers
- Hans F, Fiesel FC, Strong JC, Jäckel S, Rasse TM, Geisler S, Springer W, Schulz JB, Voigt A, Kahle PJ. UBE2E ubiquitin-conjugating enzymes and ubiquitin isopeptidase Y regulate TDP-43 protein ubiquitination. J Biol Chem. 2014 Jul 4;289(27):19164-79. Epub 2014 May 13 PubMed.
- Mackness BC, Tran MT, McClain SP, Matthews CR, Zitzewitz JA. Folding of the RNA recognition motif (RRM) domains of the amyotrophic lateral sclerosis (ALS)-linked protein TDP-43 reveals an intermediate state. J Biol Chem. 2014 Mar 21;289(12):8264-76. Epub 2014 Feb 4 PubMed.
- Jiang LL, Che MX, Zhao J, Zhou CJ, Xie MY, Li HY, He JH, Hu HY. Structural Transformation of the Amyloidogenic Core Region of TDP-43 Protein Initiates Its Aggregation and Cytoplasmic Inclusion. J Biol Chem. 2013 Jul 5;288(27):19614-24. PubMed.
- Wang YT, Kuo PH, Chiang CH, Liang JR, Chen YR, Wang S, Shen JC, Yuan HS. The Truncated C-terminal RNA Recognition Motif of TDP-43 Protein Plays a Key Role in Forming Proteinaceous Aggregates. J Biol Chem. 2013 Mar 29;288(13):9049-57. PubMed.
- Nishimoto Y, Ito D, Yagi T, Nihei Y, Tsunoda Y, Suzuki N. Characterization of alternative isoforms and inclusion body of the TAR DNA-binding protein-43. J Biol Chem. 2010 Jan 1;285(1):608-19. PubMed.
- Zhang YJ, Xu YF, Cook C, Gendron TF, Roettges P, Link CD, Lin WL, Tong J, Castanedes-Casey M, Ash P, Gass J, Rangachari V, Buratti E, Baralle F, Golde TE, Dickson DW, Petrucelli L. Aberrant cleavage of TDP-43 enhances aggregation and cellular toxicity. Proc Natl Acad Sci U S A. 2009 May 5;106(18):7607-12. PubMed.
News
- Stress Relief: Anti-Stress Granule Therapy Saves ALS Models
- Enzyme Structure Linked to ALS Severity
- Does TDP-43 Oligomerize and Coax Aβ to Do the Same?
- Good Prion, Bad Prion: TDP-43 Domain Plays Both Sides
- ALS-Linked TDP-43 Turns Amyloid in the Lab
- Double Down: TDP-43 Fragments Bust Cells on Second Hit
- Artificial Chaperone Keeps Amyloid-Forming Protein in Check
Primary Papers
- Qin H, Lim LZ, Wei Y, Song J. TDP-43 N terminus encodes a novel ubiquitin-like fold and its unfolded form in equilibrium that can be shifted by binding to ssDNA. Proc Natl Acad Sci U S A. 2014 Dec 30;111(52):18619-24. Epub 2014 Dec 12 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
ICGEB - Trieste
ICGEB-Trieste
This work by Qin and colleagues is a step toward understanding the molecular mechanisms involved in TDP-43 pathology. These results, when seen in conjunction with the recently described role of TDP-43 amino terminus region in aggregation processes that lead to TDP 43 loss of function (Budini et al., 2014), show how important the N-terminal domain is to the overall picture of ALS and FTLD pathogenesis.
Qin et al. have found that the N-terminal region of TDP-43 has a folded structure and an unfolded structure that co-exist in a tightly regulated equilibrium that depends on salt and protein concentration. This equilibrium also might be affected by intra- and intermolecular interactions. The well-folded conformation appears to assume a ubiquitin-like structure able to bind ssDNA and RNA molecules. This conformation allows TDP-43 to form oligomers, shifting the equilibrium toward the folded conformation.
These results shed light on two of the most studied features of pathogenic TDP-43—protein aggregation and loss of function. The mechanisms behind these events are poorly understood and it is still unknown if the aggregation is a cause or a consequence of the disease. The existence of an equilibrium between folded and unfolded conformations of the protein in vivo could explain several observations. For instance, in some patients affected by ALS and FTLD, increased TDP-43 levels have been found, an aspect that correlates well with the possible disturbance of the equilibrium between the folded and the unfolded states.
In pathological conditions, TDP-43 loses some of its functions. The unfolded N-terminal domain of TDP-43 may favor aggregation and at the same time hamper the stabilization of the nucleic acids at the RNA recognition motifs of the protein, leading to the impairment of TDP-43 function.
However, it remains to be seen if some other events trigger the shift of this balance. For example, environmental stress could stabilize the unfolded form. Whatever shifts the equilibrium to the unfolded structure, it will be important to understand if it is possible to intervene in the aggregation process and bring the protein back to the well-folded state, because this may prevent or repair the neuronal damage. Hence these results open new frontiers towards novel therapeutic strategies in order to prevent irreversible protein aggregation and neuronal death.
References:
Budini M, Romano V, Quadri Z, Buratti E, Baralle FE. TDP-43 loss of cellular function through aggregation requires additional structural determinants beyond its C-terminal Q/N prion-like domain. Hum Mol Genet. 2015 Jan 1;24(1):9-20. Epub 2014 Aug 13 PubMed.
View all comments by Francisco BaralleMake a Comment
To make a comment you must login or register.